Aunque la extensión de la Botánica como tema aparte, y también de la Zoología, superan con creces la intención de nuestro Foro, creemos que podemos ir subiendo artículos sobre las mismas, incluidos ya o no, en el tema Biología.
Empezamos con una visión muy interesante de la evolución de las plantas. Más adelante entregaremos definiciones de nociones y conceptos usados en Botánica.
_____________________
En:

Coevolución de las flores y sus polinizadores
Por: Ferney Yesyd Rodríguez
I. Parte
Se permite la reproducción de este ensayo para fines educacionales y/o científicos siempre y cuando se mencione claramente nuestro sitio web, así como el nombre del autor del mismo.
Se prohibe su reproducción con fines comerciales.

______________________________________________
Con frecuencia cuando se habla de evolución se nos viene a la mente los fósiles de dinosaurios o de los restos de primates extintos relacionados con los orígenes humanos, olvidando los vegetales, y otros seres, que como todas las especies, han sido moldeadas por la evolución. Por ejemplo, al leer los textos de biología para la escuela se nota que las referencias a la evolución de las plantas son casi inexistentes. Por eso, con este texto quiero llamar la atención sobre un aspecto de la evolución en el mundo vegetal: la coevolución entre las plantas con flores y sus polinizadores. Además notaremos como las adaptaciones de estos organismos fueron explicadas por Charles Darwin por medio de la selección natural.
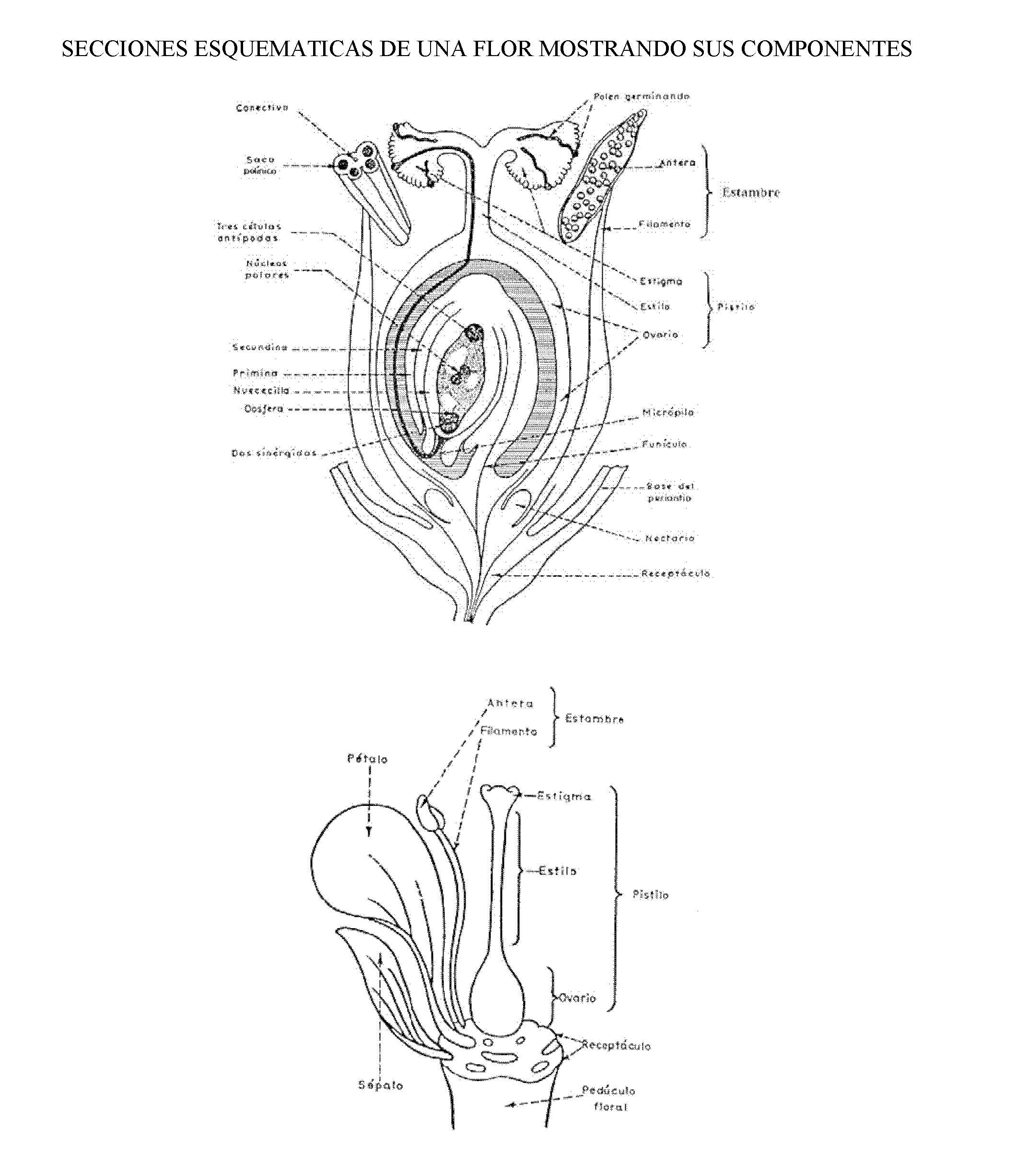
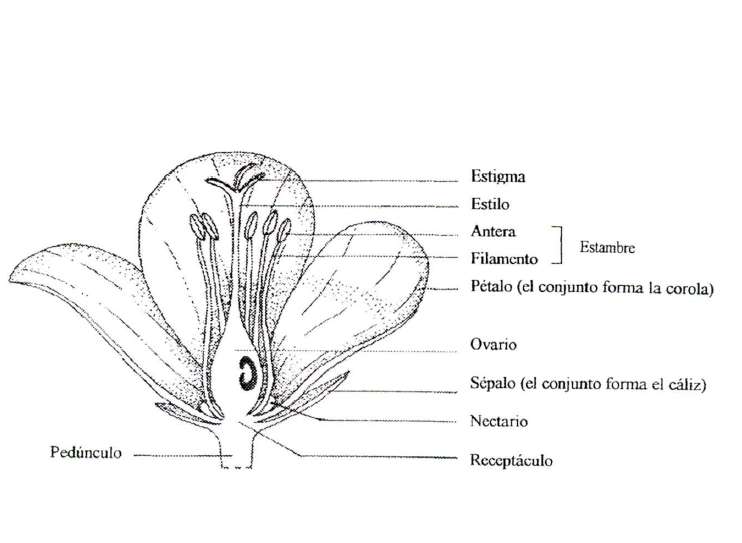
La Flor para empezar
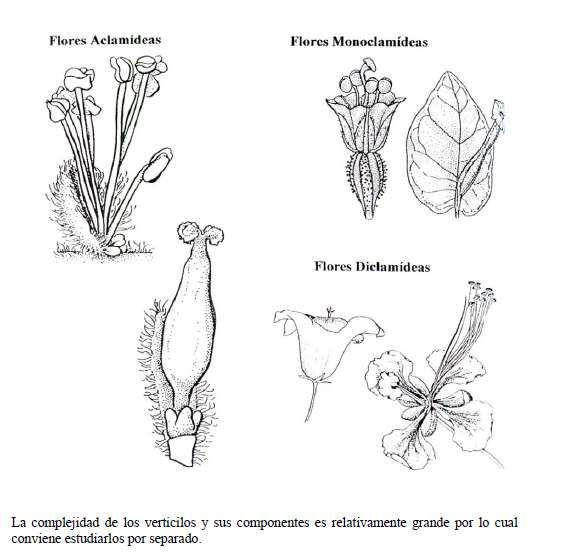
Antes de hablar de las adaptaciones de las flores a sus polinizadores es necesario recordar lo qué es una flor. Las flores son hojas modificadas que poseen tejidos reproductivos y estériles. Cada conjunto de piezas florales que nacen de un nudo reciben el nombre de verticilo y las flores poseen cuatro: dos estériles (cáliz y corola) y dos reproductores o sexuales (androceo y gineceo).
En la flor ocurren dos acontecimientos importantes en la vida de un organismo de reproducción sexual: la producción de los gametos (o células sexuales) y la fecundación. Para la fecundación es necesario el transporte desde el verticilo masculino de un grano de polen (una estructura que contiene dos células masculinas y una cubierta dura) hasta el verticilo femenino, el gineceo, para que germine y alcance el óvulo.

Las plantas con flores pueden tener autopolinización o polinización cruzada. En el primer caso el polen producido en los estambres de una flor cae en el estigma de esa misma flor. Este es el caso de las flores de la arveja, y de esta característica fue de la que se valió el famoso Gregorio Mendel para escoger a los guisantes como su organismo de experimentación en genética. En la polinización cruzada el polen producido en una flor debe ir a las estructuras femeninas de la flor de otro organismo. Con la polinización cruzada se genera mayor diversidad genética que con la autopolinización.
En las angiospermas existen muchas formas de transportar el polen, algunas plantas como los pastos dependen del viento, pero muchas otras se valen de la ayuda de animales, en estos casos hablamos de polinización biótica.

Darwin, la polinización biótica y la selección natural
Charles Darwin fue un naturalista excepcional que hizo un gran número de observaciones en el mundo vegetal. Llegó a tener un invernadero en su casa en Dowo donde hizo experimentos sobre la importancia de la fecundación, además de meticulosas observaciones que le ayudaron a sustentar la selección natural como mecanismo evolutivo. En El Origen podemos leer:
"Las flores se encuentran dentro de las producciones más hermosas de la naturaleza; pero se han vuelto visibles al contraste con las hojas verdes, y en consecuencia, hermosas al mismo tiempo, para que puedan ser vistas fácilmente por los insectos. He llegado a esta conclusión porque he encontrado como regla invariable que cuando una flor es fecundada mediante el viento, no tiene nunca una corola de color llamativo. Diversas plantas producen habitualmente dos clases de flores: Unas abiertas y coloreadas de tal modo que atraigan a los insectos, y otras cerradas, no coloreadas, desprovistas de néctar y que nunca son visitadas por los insectos. Por consiguiente podemos llegar a la conclusión de que, si los insectos no se hubiesen desarrollado sobre la faz de la Tierra, nuestras plantas no se hubieran cubierto de bellas flores y hubieran producido solamente flores tan pobres como las que vemos en el abeto, el roble, el nogal y el fresno, y en las gramíneas, espinacas, acederas, y ortigas, que se fecundan por la acción del viento." (El Origen de las Especies. Capítulo VI. Pág. 213).
"He averiguado por experimentos que las abejas y abejarrones son casi indispensables para la fertilización del pensamiento (Viola tricolor), pues otros insectos no visitan esta flor. He descubierto también que las visitas de las abejas son necesarias para la fertilización de ciertas clases de trébol; por ejemplo 20 cabezas de trébol blanco (Trifolium repens) produjeron 2290 semillas, y otras 20 cabezas resguardadas de las abejas no produjeron ninguna. Igualmente, 20 cabezas de trébol rojo (T. pratense) produjeron 2700 semillas, pero el mismo número de cabezas protegidas no produjo ninguna semilla. Solo los abejorros visitan el trébol rojo, pues los otros insectos no pueden alcanzar el néctar" (El Origen de las Especies. Capítulo IV. Pág. 110 & 111).*
"Tal vez vale la pena dar otro ejemplo más complejo de la acción de la selección natural. …supongamos que los insectos al buscar el jugo o néctar segregado desde el interior de las flores de un cierto número de plantas de una especie cualquiera quedarían empolvados de polen, y con frecuencia lo transportarían de una flor a otra; las flores de dos individuos distintos de la misma especie se cruzarían así, y el hecho del cruzamiento, como puede probarse plenamente, origina plantas vigorosas que, por tanto, tendrán más posibilidades de florecer y sobrevivir. Las plantas que produjeran glándulas o nectarios más grandes y que segregaran más néctar, serían las visitadas con mayor frecuencia por los insectos y las más frecuentemente cruzadas; y de este modo, a la larga, adquirirían ventaja y formarían una comunidad local.
Asimismo, las flores que tuvieran sus estambres y pistilos colocados en relación con el tamaño y las costumbres del insecto determinado que las visitase, de modo que facilitase en cierto modo el transporte de polen, también serían favorecidas. Pudimos haber tomado el caso de insectos que visitan flores con el objeto de recoger polen, en vez de néctar; y como el polen se produce con el único propósito de la fecundación su destrucción parece ser una simple perdida para la planta; sin embargo, si un poco de polen fuese llevado de una flor a otra, primero accidentalmente, y luego habitualmente, por los insectos que devoran polen, efectuándose de este modo un cruzamiento, aunque los nueve décimas partes fueran destruidas, todavía podría ser un gran beneficio para la planta ser robada así, y los individuos que produjeran cada vez más polen y anteras más grandes serían seleccionados.*
"tan pronto como la planta se hubiese vuelto tan atractiva para los insectos que el polen era llevado regularmente de flor en flor, pudo comenzar otro proceso. Ningún naturalista duda de la división fisiológica del trabajo; por consiguiente podríamos creer que sería ventajoso para una planta producir estambres solos en una flor o en toda una planta, y pistilos solo en una flor o en toda una planta.
Volvamos ahora a los insectos que se alimentan de néctar; podemos suponer que la planta cuyo néctar hemos hecho aumentar lentamente por selección continuada, sea una planta común, y que ciertos insectos dependan principalmente de su néctar para alimentarse. Podría citar muchos hechos para demostrar lo codiciosas que son las abejas para ahorrar tiempo; por ejemplo su costumbre para hacer agujeros y de chupar el néctar en la base de ciertas flores
Teniendo en cuenta estos hechos, puede creerse que, en ciertas circunstancias, diferencias individuales en la curvatura o longitud de la trompa, etc. demasiado ligeras para ser apreciadas por nosotros, podría aprovecharlas una abeja u otro insecto, de hecho que ciertos individuos fueran capaces de alcanzar su alimento más rápidamente que otros; y así las comunidades a que ellos perteneciesen prosperarían y dejarían muchos enjambres que heredarían las mismas particularidades. Los tubos de la corola del trébol rojo común y del encarnado (*Trifolium platense y T. incarnatum) no parecen diferir a simple vista, en longitud; sin embargo, la abeja común la abeja común puede chupar fácilmente el néctar del trébol encarnado, pero no el del trébol rojo, que es visitado por los abejorros; de modo que campos enteros de trébol rojo ofrecen en vano una abundante provisión de precioso néctar a la abeja común… Por otra parte, como la fecundidad de éste trébol depende por completo de las abejas que visitan las flores, si los abejorros llegasen a ser raros en algún país, podría ser una gran ventaja para la planta tener una corola más corta o más profundamente separada, de suerte que otro insecto pudiese succionar sus flores. Así puedo comprender como una flor y una abeja pudieron lentamente, y de una manera simultánea o una después de otra, modificarse y adaptarse entre si del modo más perfecto, mediante la conservación continuada de todos los individuos que presentasen ligeras desviaciones de estructura mutuamente favorables."* (El Origen de las Especies. Capítulo IV. Pág. 126-128. Énfasis añadido.)
Como lo notaba Darwin, la belleza de las flores ha evolucionado para permitir la fecundación cruzada. Esta polinización puede ser llevada a cabo por muchas especies animales, y en la interacción entre las plantas y sus polinizadores, la selección natural ha generado una gran variedad de adaptaciones en las plantas para garantizar que sus granos de polen lleguen a estigmas de otras flores de su misma especie. Las características de las flores que atraen a los polinizadores pueden clasificarse como señales y recompensas.
Las señales avisan a los polinizadores que hay alimento presente en la flor y cuando el animal visita la flor le es transferido el polen permitiendo que este alcance otra flor. Los tipos de señales presentadas por las flores a los animales varían entre las diferentes especies vegetales. Estas pueden ser visuales (color, textura y patrón), olfatorias (esencias) o movimiento, en algunos casos aún. No es poco probable que haya más de una señal presente en una flor.
Las recompensas ofrecidas por las plantas a los polinizadores casi siempre involucran una fuente de alimento, la más común de esta se denomina néctar. El néctar es un complejo de azucares, aminoácidos y compuestos aromáticos. Usualmente el néctar no es lo que da el aroma a las flores (el aroma es una señal y el néctar es una recompensa). Algunas plantas ofrecen ceras como alimento, mientras que algunas veces el polen mismo es usado para tal fin.
Es importante señalar que la mayoría de las especies de plantas no son polinizadas por un tipo determinado de polinizador.
Sin embargo, hay casos en los cuales una especie particular poliniza una planta (en algunas orquídeas), pero estos casos son la excepción.
Para la mayoría de las especies de plantas se pueden hacer generalizaciones sobre las señales que presentan las flores que son polinizadas por grupos particulares de animales como insectos (escarabajos, mariposas, polillas, moscas, abejas, avispas), aves y murciélagos. Estas señales particulares, que son adaptaciones a un tipo de polinizador particular, aparecieron como resultado de la selección natural y hacen parte de un proceso de coevolución.
La coevolución de las plantas con flores
La coevolución es una serie de cambios evolutivos que se dan de forma reciproca entre dos o más especies ecológicamente interrelacionadas.
Para que se presente la coevolución se requiere que se de una serie de cambios en las especies implicadas. No se trata simplemente que una especie presente un cambio como resultado de la actividad o la presencia de otro organismo. Para que haya coevolución es necesario las adaptaciones que desarrolla la especie 1 sea resultado de las adaptaciones de la especie 2 y de esta manera la especie 1 pueda incrementar su éxito reproductivo (o "fitness"), luego la especie 2 desarrollará por selección natural otra adaptación (o una mejora de las ya presentes) que le permitirán utilizar las características de la especie 1 para dejar más descendencia. Esta influencia evolutiva mutua llevará a establecer una relación de mutualismo entre las especies 1 y 2.

Como señalara Darwin, "la selección natural probablemente no puede producir ninguna modificación de una especie exclusivamente para el bien de otra especie; aunque por toda la naturaleza unas especies sacan incesantes ventajas y provecho de las estructuras de otras". (El Origen de las Especies. Capítulo VI. Pág. 214) Las plantas que producen recompensas a los polinizadores no lo hacen porque quieran alimentarlos gratuitamente, esta característica evolucionó porque asegura la llegada del polen a otras flores de su misma especie y de esta manera pueden dejar más descendencia. Las plantas que no invierten en recompensas, como los pastos deben producir una cantidad mucho mayor de polen, pues dependen del viento para la polinización cruzada. De igualmente manera, los polinizadores no visitan las flores porque estén interesados en ayudar a las plantas a reproducirse, ellos simplemente van en busca de una fuente de energía que les permite seguir viviendo y eventualmente reproducirse y dejar descendencia también.

Algunos casos de coevolución han llevado a que solo una especie de polinizador pueda acceder al néctar de una especie vegetal, por lo que esta última terminará dependiendo de la existencia de este polinizador para hacer llegar su polen a los estigmas de otra flor de la misma especie. Este caso se presenta en la orquídea estrella de navidad (Angraecum sesquipedale), la cual se caracteriza por tener un estrecho recipiente tubular, llamado espolón (presente en muchas especies de orquídeas), que contiene el néctar. El espolón de la orquídea estrella de Navidad es inmenso, comparado con el de sus congéneres, pues mide 29 centímetros de longitud. En el fondo de este recipiente, el néctar no alcanza a ocupar más de cuatro centímetros.
La orquídea estrella de navidad fue descrita por el botánico francés Louis-Marie-Aubert du Petit-Thouars quien trabajó en Madagascar y otras regiones de África a finales del siglo XVIII e inicios del XIX. Sin embargo, él botánico francés no conoció un polinizador que pudiera acceder al néctar ubicado tan profundamente. En 1862, Charles Darwin en una obra sobre la fertilización de las orquídeas ("On the various contrivances by wich British and foreign orchids are fertilized by insects, and on the good effects of intercrossing"), propuso una solución al enigma: El polinizador de la particular orquídea malgache debía ser una mariposa con una espiritrompa con una longitud entre 25 y 28 centímetros. Como por aquella época no se conocía ninguna mariposa con una trompa tan larga, varios entomólogos ridiculizaron la hipótesis de Darwin.
No fue sino hasta 1910 que el entomólogo Karl Jordan y Lionel Walter Rothschild encontraron el insecto que polinizaba la orquídea estrella de navidad. El insecto era una subespecie (raza geográfica) de la esfinge de Morgán (Xanthopan morgani preadicta). Sin embargo, en 1873 el codescubridor del mecanismo de la selección natural, el naturalista Alfred Russel Wallace, ya había sugerido como polinizador a la esfinge de Morgan, la cual en el continente africano tiene una espiritrompa de unos 20 centímetros de longitud.
_________________________________