___________________________________________________
Es deseable que la biologia, y la ciencia en general, la pensemos como sistemas, con una concepcion tactica y estrategica a la vez, vale decir, a corto y largo plazo. Esta introduccion y todo el libro, es un formidable ejemplo que envuelve conceptos y principios generalizadores de los seres vivos, en este caso, de los vertebrados.____________________________________________________
Anatomia Comparada (Vertebrados)
Alfred S. RomerEditorial Interamericana S.A. 1966____________________________________________________________
INTRODUCCION
Esta obra intenta presentar en forma abreviada la historia del cuerpo de los vertebrados. Es fundamental el estudio comparativo de la estructura de los vertebrados, o sea, la anatomía comparada; es una disciplina interesante y de gran provecho. Sin embargo, tiene mayores alcances el hecho de que las modificaciones estructurales que explicaremos implican cambios funcionales experimentados por los vertebrados, cambios que, a su vez, dependen de la diversidad del ambiente y los modos de vida en el curso de su larga historia rica en acontecimientos. La evolución de los vertebrados se conoce mejor que la de cualquier otro grupo de animales, y brinda magnífica ilustración de muchos principios biológicos generales. Conocer la estructura de los vertebrados tiene valor práctico para quienes se dedican a muchos campos de la biología animal. Al futuro estudiante de medicina le brindará una mejor comprensión de la especie animal que habrá de estudiar más tarde.
En su mayor parte (capítulos 6 a 17) esta obra se dedica a explicar, de manera seriada, los diversos órganos y sistemas de órganos. En este capítulo daremos una noción a vuelo de pájaro de la estructura de los vertebrados, y presentaremos algunos temas preliminares. En otros capítulos iniciales trataremos temas generales o preliminares, incluyendo la historia evolutiva de los vertebrados y sus semejantes (caps. 2 y 3); células y tejidos como elementos estructurales básicos (cap. 4), y desarrollo embrionario (cap. 5). PLAN DEL CUERPO DE LOS VERTEBRADOS Simetría bilateral. Un carácter básico de la norma estructural de los vertebrados es que los miembros de este grupo presentan simetría bilateral: un lado del cuerpo es, en esencia, imagen en espejo del otro. Los vertebrados comparten esta clase de organización con diversos grupos de invertebrados, principalmente los anélidos y el extenso filo de los artrópodos, que incluye crustáceos, arácnidos, insectos, etc., En cambio, encontramos simetría radial en celentéreos y equinodermos, en los cuales las diversas partes de la economía nacen de una porción central desde la cual se proyectan como los rayos de una rueda. El grado de actividad de los animales parece guardar relación con el tipo de simetría que poseen. Los equinodermos y celentéreos radiales son, en términos generales, poco activos, de movimientos lentos o fijos al fondo; si flotan libremente, se dejan transportar por la corriente y no nadan activamente. Por otra parte, los vertebrados, artrópodos y anélidos marinos suelen mostrar más actividad. Esta parece haber sido una de las claves del éxito de los vertebrados y, en parte, es tan característica como cualquier rasgo anatómico.
Diferenciación regional. En todo animal con simetría bilateral se observa alguna clase de división longitudinal en regiones sucesivas; por ejemplo, en los gusanos anélidos hay una repetición bastante monótona de segmentos prácticamente iguales; en los insectos los segmentos se han fusionado en cabeza, tórax y abdomen. También los vertebrados poseen regiones corporales bien definidas, aunque no pueden compararse directamente con las de los invertebrados.
En los vertebrados encontramos una región cefálica o cabeza muy especializada, en ella se concentran los principales órganos de los sentidos, los centros nerviosos de más importancia que forman el encéfalo, y la boca y estructuras relacionadas con ella. En este caso, como ocurre en todos los animales con simetría bilateral (incluso los gusanos) se advierte tendencia notable hacia la cefalización, que consiste en la concentración de estructuras y funciones en el extremo anterior del cuerpo.
En todos los vertebrados superiores terrestres, existe un cuello detrás de la cabeza; es poco más que un segmento de unión que permite movimientos de la cabeza sobre el tronco. Sin embargo, la presencia de cuello no es un carácter fundamental de los vertebrados. En los vertebrados inferiores que viven en el agua, este segmento del cuerpo es la región branquial, corta y gruesa, que contiene el aparato respiratorio. El cuello se presenta de manera neta con el paso a la respiración pulmonar, con disminución de las branquias.
La porción más grande del animal, el tronco, es la región que sigue; termina cerca del ano o de la cloaca, En el tronco están las cavidades corporales, que contienen órganos muy importantes, las vísceras. En los mamíferos el tronco se divide en tórax y abdomen; el primero contiene corazón y pulmones dentro de la caja ósea formada por las costillas; el segundo incluye la mayor parte del aparato digestivo; sin embargo, en los vertebrados inferiores no hay una subdivisión neta.
En la mayor parte de invertebrados con simetría bilateral, el tubo digestivo abarca casi toda la longitud del cuerpo. Sin embargo, entre los vertebrados el aparato digestivo y otras vísceras distan bastante de llegar al extremo del cuerpo; detrás del tronco es característico que se extienda la cola o región caudal, bien desarrollada, con músculos y huesos, pero sin vísceras. Claro está, la cola es el principal órgano propulsor en los vertebrados primitivos acuáticos. En los animales terrestres tiende a perder importancia, pero a menudo es larga y gruesa en la base, y está bien desarrollada en muchos anfibios y reptiles. Suele persistir en los mamíferos pero es solamente un apéndice delgado. En las aves ha experimentado acortamiento y substitución funcional por las plumas de la cola, que nacen del muñón caudal; en algunas especies ranas, primates y hombre se observa el hecho excepcional de su completa desaparición como estructura externa.
Branquias. Uno de los caracteres más peculiares (quizá el más distintivo) de los vertebrados y formas vecinas es la presencia, si no en el adulto en el embrión, de branquias internas, bajo forma de una serie pareada de hendiduras o sacos que se dirigen al exterior desde la porción anterior del intestino, o sea la faringe. En los vertebrados superiores las branquias han sido substituidas por pulmones, pero en el embrión siguen existiendo sacos branquiales. En los vertebrados inferiores acuáticos los órganos principales de la respiración son las branquias. Muchos invertebrados pequeños de superficie membranosa y blanda pueden obtener a través de la misma oxígeno suficiente para satisfacer sus necesidades, Sin embargo, los animales de superficie dura o en forma de concha (sobre todo los grandes, en los cuales el área superficial es pequeña comparada con el volumen del cuerpo) necesitan estructuras branquiales de alguna índole. Las branquias características de los invertebrados, por ejemplo las de crustáceos o moluscos, son prolongaciones plumosas de la superficie corporal. Sin embargo, las branquias de los vertebrados son órganos internos conectados con el tubo digestivo. El agua entra a la garganta o faringe (generalmente por la boca) y pasa por hendiduras o sacos, en la superficie de estas vías hay membranas branquiales, en las cuales ocurre el intercambio de oxígeno del agua por bióxido carbónico de la sangre. Contrasta mucho con lo anterior la función de las branquias en algunos familiares inferiores de los vertebrados; como después veremos, las branquias y hendiduras branquiales son de importancia primordial para la obtención de alimentos; este hecho quizá explique la circunstancia insólita de que en los vertebrados los órganos de la respiración estén relacionados con el tubo digestivo.
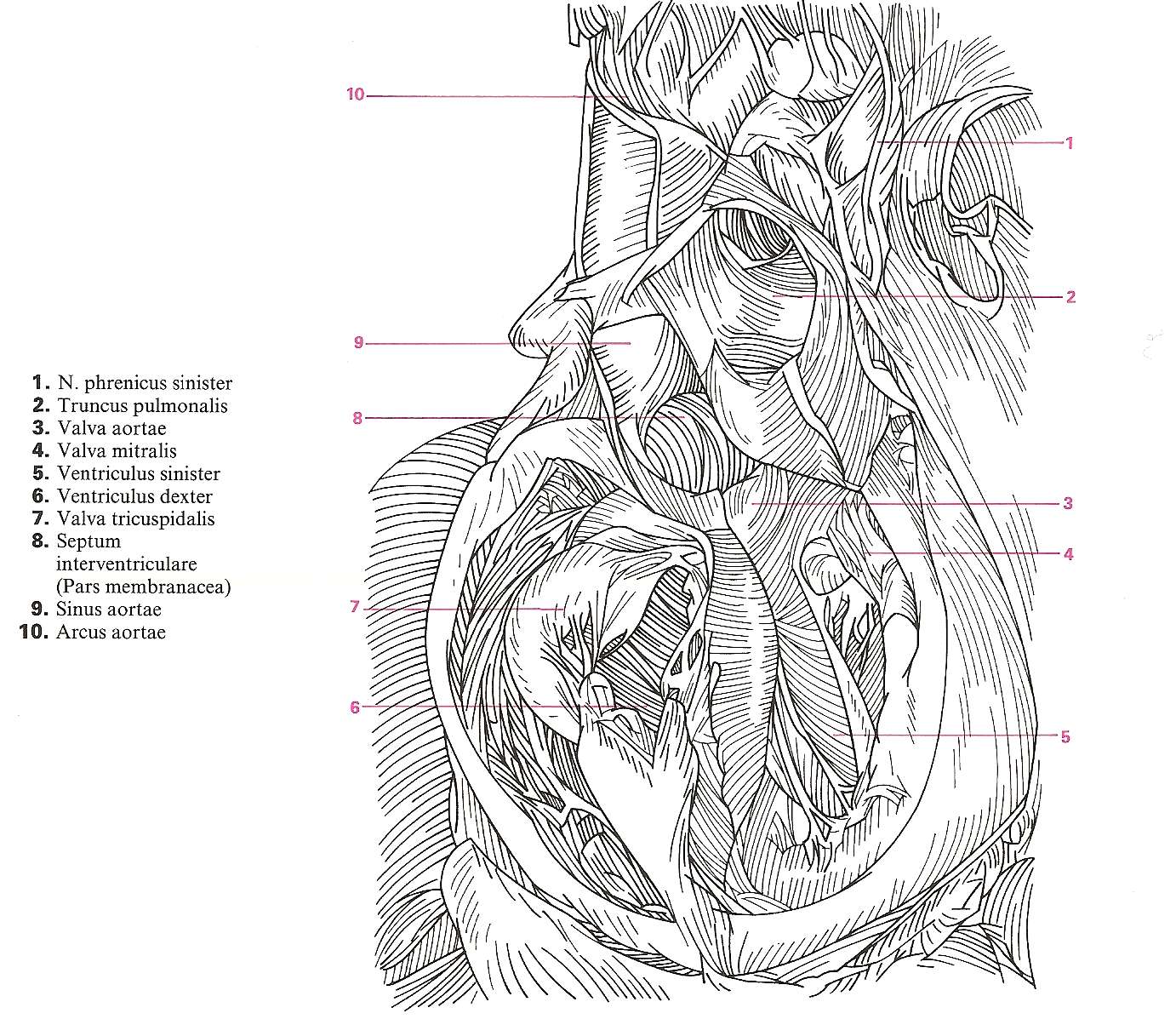
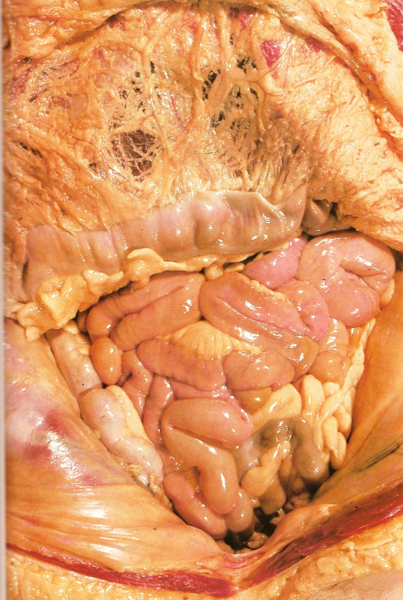
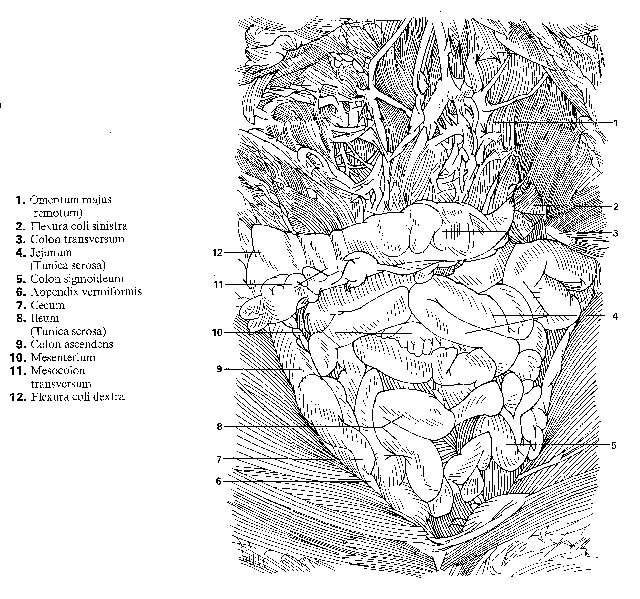
FIG. 1. Esquema de un corte longitudinal de un vertebrado ideal donde se observa la posición de los principales órganos. Notocordio.
Notocordio. En el embrión de todos los vertebrados se advierte una estructura larga, flexible y cilíndrica que se extiende de la cabeza a la cola a lo largo del dorso; es el notocordio. En la mayor parte de vertebrados el notocordio es rudimentario o falta por completo en el adulto, pues ha sido substituido por la columna vertebral. Sin embargo, sigue siendo notable en algunos vertebrados inferiores, y es el sostén principal del tronco en algunas formas de constitución sencilla (por ejemplo Amphioxus) en las cuales nunca llega a formarse columna vertebral. Es tan importante esta estructura primitiva de sostén, que los vertebrados constituyen el filo de los cordados (Chordata), nombre que denota que hay notocordio.
Sistema nervioso. En varios grupos de invertebrados de simetría bilateral, existen cordones nerviosos longitudinales; sin embargo, a menudo son pareados y pueden tener posición lateral o ventral. Sólo en los cordados se advierte un cordón único de situación dorsal dispuesto a lo largo del dorso por encima del notocordio o las vértebras. Los cordones nerviosos de los invertebrados son masas compactas de fibras nerviosas (y células de sostén) dispuestas entre cúmulos también macizos de neuronas, llamados ganglios. En cambio, el cordón neural de los cordados es una estructura hueca, sin ganglios, que posee una cavidad central llena de líquido. En varios invertebrados el fenómeno de cefalización hace que se concentren los centros nerviosos en una estructura encefaloide. Independientemente, a nuestro juicio, los vertebrados han desarrollado un cerebro hueco, con subdivisiones características, en el extremo anterior del tubo neural o cordón neural hueco (médula espinal). Tampoco se halla equivalente exacto en ningún grupo de invertebrados de una serie de órganos de los sentidos característicos que se desarrollan en la región cefálica de los vertebrados: ojos pareados y, antiguamente, un tercer ojo central, estructuras nasales, por lo regular pareadas; oídos pareados cuya función principal es el equilibrio.
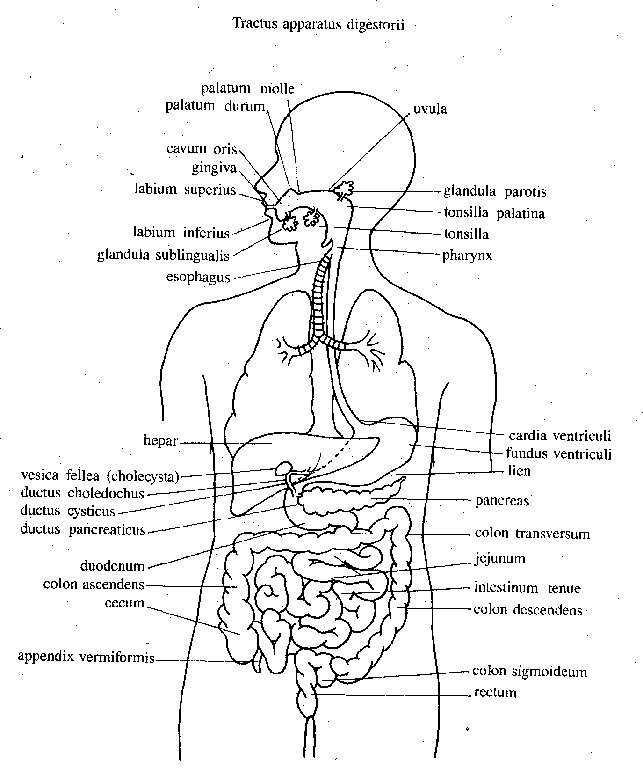
Aparato digestivo. Todos los metazoarios (con excepción de los que han experimentado degeneración) presentan alguna clase de cavidad digestiva con entrada y salida. En muchos metazoarios muy primitivos sólo hay un orificio, que funciona como boca y ano. En los vertebrados, igual que en otros metazoarios evolucionados, hay orificios anterior y posterior separados. La boca está situada cerca del extremo cefálico o anterior del cuerpo, y de manera característica algo hacia abajo. En artrópodos y anélidos el tubo digestivo llega al extremo posterior del cuerpo. No así en los vertebrados, donde el ano está situado en el extremo del tronco, y queda la región caudal en la que no hay tubo digestivo, según mencionamos.
En la mayor parte de vertebrados el tubo digestivo se divide en regiones características con funciones variadas: boca, faringe, esófago, estómago e intestino (este último se subdivide de manera variable). En los vertebrados inferiores el esófago puede faltar casi por completo, y en algunos grupos el estómago también. En los mamíferos y otros vertebrados el aparato digestivo termina en el ano, Sin embargo, casi en todos los grupos hay un segmento intestinal terminal, la cloaca, donde desembocan también los conductos urinarios y genitales.
El tubo digestivo de los vertebrados presenta una excrescencia ventral voluminosa y única, el hígado, que tiene en cierta medida función secretora, pero que principalmente actúa como órgano de almacenamiento y transformación de los alimentos. En muchos invertebrados hay vísceras algo semejantes pero de carácter variable.
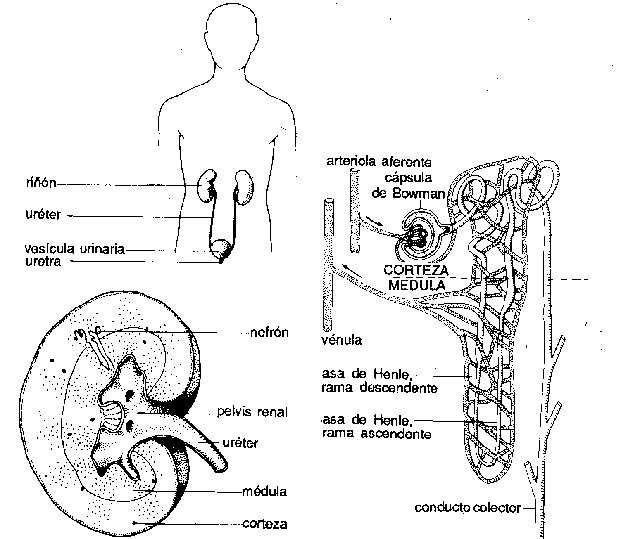
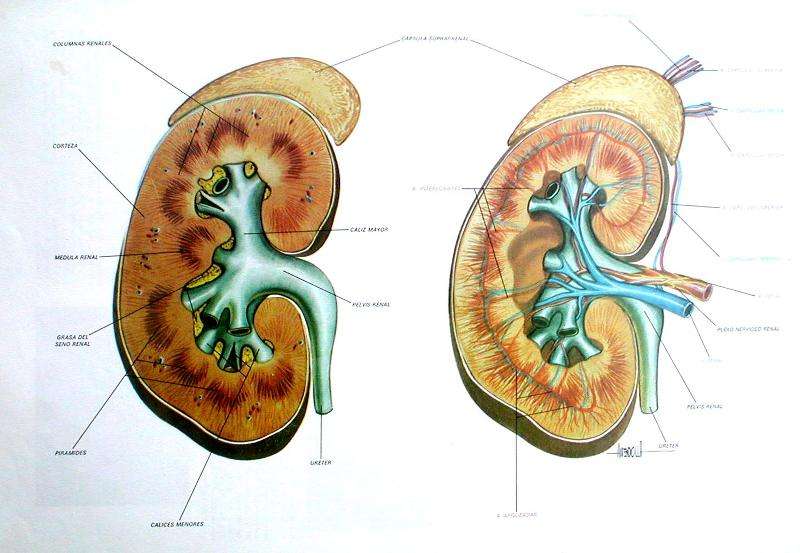
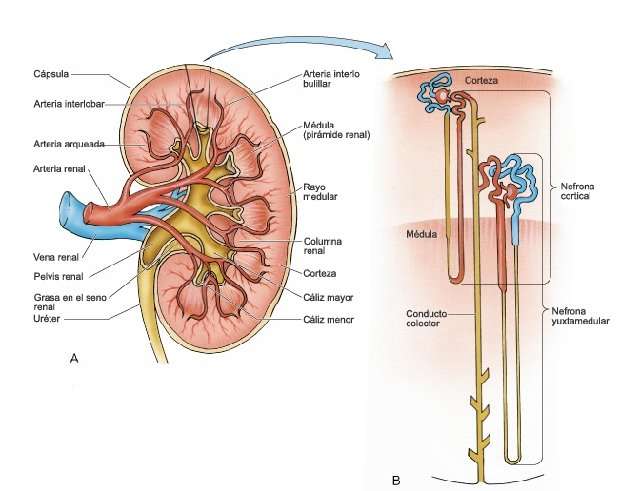
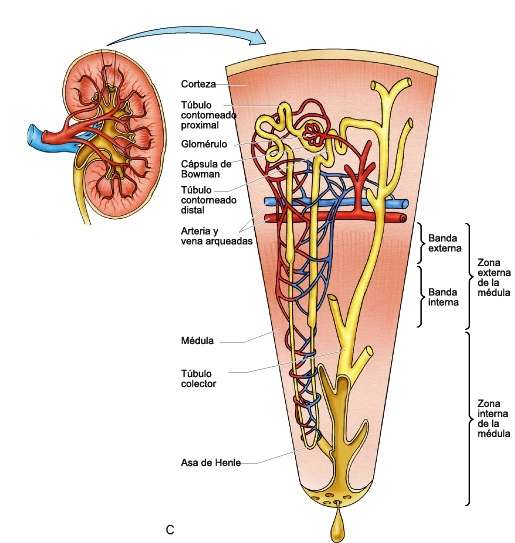
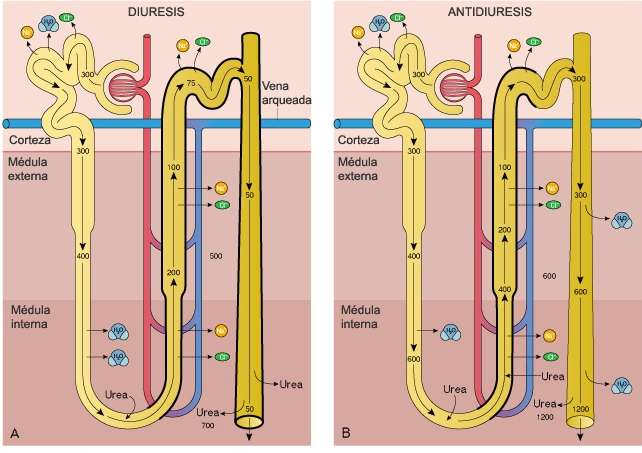
Riñones. Los invertebrados a menudo poseen algún órgano para la eliminación de desechos nitrogenados y para mantener la composición adecuada de los líquidos internos de la economía; muchas veces, se trata de hileras de pequeñas estructuras tubulares llamadas nefridios. Entre los cordados inferiores, Amphioxus posee nefridios de tipo especial. En los vertebrados verdaderos los túbulos renales que desempeñan, la función mencionada difieren mucho en tipo y suelen agruparse en órganos pares y compactos de situación dorsal. Los conductos renales, que varían en características, desembocan en la cloaca o en el exterior, y en su trayecto puede desarrollarse una vejiga urinaria.
Órganos de la reproducción. En los vertebrados casi siempre hay separación neta de machos y hembras, igual que en muchos grupos de invertebrados. Los tejidos que poseen las células germinativas las gónadas se convierten en testículo u ovario. En todos los vertebrados, excepto en los más inferiores, hay un sistema de conductos que lleva los óvulos o espermatozoides hacia la superficie, o de la superficie al interior (a menudo por la cloaca); en la hembra, puede haber formaciones especiales para la producción de una envoltura resistente (o cáscara) del huevo, para facilitar el desarrollo del embrión.
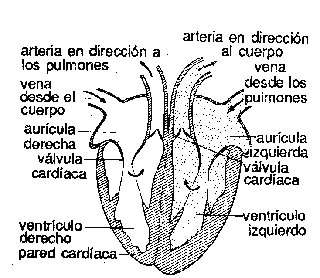
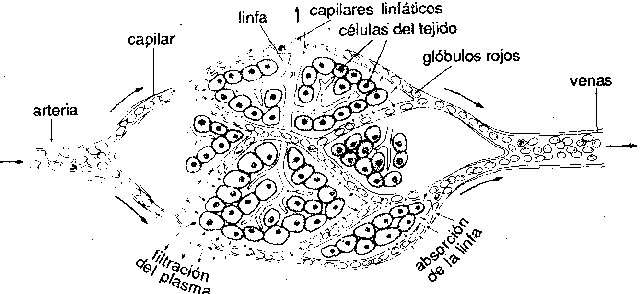
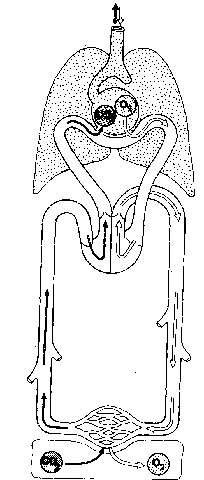
Sistema circulatorio. En los vertebrados, como en muchos invertebrados, hay un sistema orgánico bien desarrollado que contiene un líquido corporal, la sangre, con vasos tubulares y una bomba, el corazón, para hacer circular el líquido. En los vertebrados, el corazón es una estructura unitaria de situación ventral y bastante anterior. En algunos invertebrados la circulación es de tipo abierto. El corazón bombea sangre a los tejidos en vasos cerrados, pero después la sangre se pone en libertad y vuelve al corazón rezumando por los tejidos sin cursar por vasos. En los vertebrados y en algunos invertebrados superiores, el sistema es cerrado; la sangre es transportada a los órganos por arterias; después de pasar por los tejidos en tubos de pequeño calibre, los capilares, vuelve al corazón también en vasos cerrados, las venas. En la mayor parte de vertebrados hay vasos linfáticos, que son un camino adicional para el retorno de líquido de las células hacia el corazón. Muchos invertebrados presentan en la corriente sanguínea compuestos metálicos pigmentados en disolución, que ayudan a transportar oxígeno. En lo que se refiere a los vertebrados, el compuesto férrico hemoglobina es el portador de oxígeno casi siempre; además, no está en libertad en la sangre, sino encerrado en glóbulos rojos.
En los anélidos la circulación de la sangre es, en términos generales, anterógrada por la porción dorsal del cuerpo, y retorna ventralmente al volver de los tejidos. Lo inverso ocurre en los vertebrados. La sangre del corazón se dirige hacia adelante y arriba (antiguamente por las branquias) y luego hacia atrás para alcanzar los órganos del tronco y la cola, y la principal vía de retorno que proviene del aparato digestivo tiene situación ventral en relación con el intestino (aunque son importantes las venas dorsales).
Celoma. En algunos invertebrados los órganos internos están incluidos en los tejidos corporales. Sin embargo, en otros existen cavidades corporales o celomas llenas de líquido acuoso, en las cuales están la mayor parte de los órganos. Esto último se observa en los vertebrados. Una cavidad corporal importante, la cavidad abdominal, ocupa gran parte del tronco y contiene casi todo el aparato digestivo; en ellas sobresalen otros órganos (de la reproducción, urinario). En la porción anterior se advierte la cavidad pericárdica discreta que envuelve al corazón, y en los mamíferos los pulmones ocupan cavidades pleurales separadas.
Músculos. Los músculos de los vertebrados son de dos clases: estriados y lisos (no estriados), que difieren notablemente por su estructura histológica y distribución. En términos generales, los músculos estriados incluyen todos los músculos voluntarios de cabeza, tronco, miembros y cola, y los músculos de la región branquial; los músculos lisos, más difundidos, se encuentran principalmente en el aparato digestivo. El músculo cardiaco posee, en varios aspectos, estructura histológica intermedia. A diferencia de la mayor parte de los demás sistemas, los músculos estriados del tronco se desarrollan en forma de una serie de unidades segmentarias.
Esqueleto. En todos los vertebrados hay substancias esqueléticas duras; siempre son óseas, en parte cuando menos (salvo en algunos grupos degenerados o quizá más antiguos). Las porciones esqueléticas superficiales (esqueleto dérmico) equivalen funcionalmente al caparazón de algunos invertebrados, y son característicamente óseas; las estructuras esqueléticas internas (endoesqueleto) se forman como cartílago en el embrión, pero a menudo se transforman en hueso en el adulto. En los invertebrados se advierten substancias cartilaginosas. Sin embargo, el hueso es peculiar de los vertebrados.
Difiere, además, en textura e histología de los materiales esqueléticos quitinosos o calcáreos de los invertebrados; asimismo, las sales que se depositan en estos tejidos son principalmente fosfato cálcico; ésta es una diferencia con la mayor parte de las estructuras esqueléticas de los invertebrados (aunque no con todas), en las cuales el compuesto cálcico corriente es el carbonato.
Miembros o apéndices. En la mayor parte de vertebrados hay dos pares de extremidades, pectorales y pélvicas, en forma de aletas o patas, que adquieren carácter cada vez más notable en los animales superiores del grupo. Sin embargo, están poco desarrolladas o faltan en los vertebrados inferiores, vivientes y extintos, de manera que no son por completo peculiares. Su estructura (a diferencia de los apéndices de los artrópodos) incluye elementos esqueléticos internos, donde se insertan músculos que se encargan del movimiento.
Segmentación. Entre los invertebrados, los artrópodos y anélidos se caracterizan por su metamerización: es la repetición seriada de partes del cuerpo en una gran serie de segmentos corporales. En los anélidos es patente la segmentación; en los artrópodos la estructura metamérica puede ser menos evidente en el adulto, pero se advierte con claridad en embriones o larvas.
Los vertebrados también son segmentados, pero la segmentación es limitada, y se ha desarrollado independientemente de la que presentan los invertebrados, en los cuales todas las estructuras, desde la piel hasta el intestino, presentan segmentación. En los vertebrados no hay segmentación de la piel ni del intestino; la disposición metamérica se advierte principalmente en los músculos del tronco. Sin embargo, en relación con las inserciones de estos músculos y con su inervación, gran parte del esqueleto y del sistema nervioso han adquirido carácter segmentario.
Cuerpo de un vertebrado estudiado en corte (fig. 2). Hemos señalado algunos de los caracteres corporales más importantes, considerando en especial muchas veces su posición anterior o posterior en el cuerpo. Ahora conviene referirnos brevemente a la organización general del cuerpo observada en cortes transversales.
FIG. 2. Cortes transversales del cuerpo de un vertebrado. A, B, Cortes muy simplificados de la cola y el tronco; se observa la estructura básica del tronco en forma de tubo doble; falta en la cola el tubo interno del intestino. C, D, Cortes esquemáticos con mayor detalle de la cola y el tronco, en los cuales se observa la posición característica de los principales órganos. 
Desde el punto de vista estructural, la región más sencilla del cuerpo es la cola, muy desarrollada en muchos grupos de vertebrados. El corte característico de la cola (fig. 2, A, C) es un óvalo alargado, con la superficie cubierta de piel. Algo por arriba del centro se advierten el notocordio (o la región central de las vértebras que lo substituye de manera característica en el adulto); hacia arriba, el tubo neural; ambas estructuras guardan invariablemente íntima relación topográfica. La cavidad corporal y las vísceras relacionadas no existen en la región caudal; las representan (en cierto sentido) los vasos sanguíneos caudales situados debajo del notocordio. Casi todo el resto de la cola está ocupado por músculos, generalmente potentes. Los músculos están dispuestos en mitades derecha e izquierda, con un tabique mediano divisor arriba y abajo.
Un corte transversal característico por el tronco es más complicado, incluso cuando se presenta de manera muy general, como se observa en la figura 2, B. Cabe considerar que el tronco es, en esencia, un sistema tubular doble, cuya estructura podría compararse en general a la cubierta y la cámara de un neumático de automóvil. El tubo exterior incluye todos los elementos principales observados en el corte transversal de la cola: notocordio, tubo neural y músculos que descienden en ambos lados debajo del revestimiento externo de piel. Hacia adentro, parecería que la pequeña zona situada debajo del notocordio en la cola, que sólo incluía vasos sanguíneos, se hubiese dilatado hasta alcanzar proporciones enormes y formar la porción abdominal del celoma. Al desarrollarse esta cavidad, el tubo exterior del tronco adquiere superficie interna además de la externa. La superficie que tapiza la cavidad corporal es el peritoneo, y la porción del mismo que forma la superficie interna del tubo externo es el peritoneo parietal. La porción del tubo externo situada entre el celoma y la superficie del cuerpo es la pared corporal.
El tubo interno constituye de manera principal el aparato digestivo. El revestimiento externo, orientado hacia el celoma, es el peritoneo visceral. El revestimiento interno es el epitelio que tapiza el aparato digestivo. Entre ellos, de manera análoga a los músculos de la pared corporal, se advierten músculo liso y tejido conectivo. En el embrión el intestino está conectado con el tubo externo en las porciones dorsal y ventral por mesenterios, que son láminas delgadas cubiertas en ambos lados por peritoneo. El mesenterio dorsal, situado arriba del intestino, siempre persiste, pero la porción ventral a menudo desaparece casi por completo.
Más adelante explicaremos en detalle la disposición de los órganos en el celoma; sin embargo, conviene ampliar algo la explicación de la situación de las vísceras. (Debe señalarse que el tamaño comparativo de la cavidad corporal nunca es tan grande como se representa en este esquema y en otros; en realidad, las vísceras llenan la mayor parte del espacio disponible.) En la figura 2, D, se ve que el tubo digestivo no es tal tubo simplemente; posee varias excrecencias, las más características, siendo el hígado, hacia abajo, y el páncreas, hacia arriba. Trátase de estructuras mediales (aunque pueden ocupar otra situación en el adulto, siempre son centrales en el embrión) y se desarrollan dentro de los mesenterios ventral y dorsal. Además, cabe que otros órganos sobresalgan en la cavidad corporal, pero proceden de tejidos situados fuera de la misma. En muchos grupos de animales los riñones sobresalen en la cavidad al abdominal en los bordes superoexternos, y es característico que los órganos de la reproducción ovarios o testículos sobresalgan en la cavidad más medialmente siguiendo su borde superior.
DIRECCIONES Y PLANOS (Capitulo dado mas arriba)
El cuerpo de los vertebrados es, en esencia, una estructura con simetría bilateral; sin embargo, hay muchas excepciones de esta afirmación general. Los órganos que en etapa primitiva estaban situados en la línea media pueden ser desplazados; así ocurre, por ejemplo, con el corazón; la porción abdominal del aparato digestivo estómago e intestino suele experimentar torsión y el intestino en ocasiones presenta una disposición asimétrica complicada. Asimismo, en órganos pares puede haber grandes diferencias; por ejemplo, en las aves adultas sólo un ovario (el izquierdo) es funcional. En el lenguado se advierte asimetría todavía mayor, pues toda la forma del cuerpo está modificada porque los lados del animal se tornan superior e inferior.
En la teoría o en la práctica el cuerpo de un animal puede ser seccionado de diversas maneras y en distintos ángulos. Si se considera que el cuerpo es cortado de través, como se haría con un salchichón, el plano de corte se considera transversal. Si la línea de corte es vertical y longitudinal, del hocico a la cola, el plano es sagital. En ocasiones este nombre se circunscribe a los cortes precisamente por la línea media el plano sagital medio y los cortes semejantes paralelos se denominan parasagitales,
Sin embargo, a menudo estos cortes se consideran partes de una serie de cortes sagitales en sentido general. El tercer plano principal de sección, en la dirección restante, consiste en secciones que atraviesan toda la longitud del cuerpo, pero horizontalmente, y abarcando toda la anchura del cuerpo. Este plano se llama frontal (paralelo a la frente del animal).
Es importante fijar una terminología direccional para describir relaciones estructurales y dar nombre a diversos órganos. Explicaremos los términos de este grupo, que fijan una posición o señalan dirección.
En la mayor parte de los vertebrados, los extremos cefálico y caudal indican la dirección hacia la cual normalmente se mueve el animal. En este sentido, son nombres corrientes de posición anterior y posterior; se usan menos craneal y caudal, pero son sinónimos, en esencia. Es lógico que las superficies superior e inferior del dorso y del vientre se llamen dorsal y ventral. La fijación en el plano transversal se relaciona con la línea media; medial o interno denota posición hacia la línea media; lateral o externo un sitio más alejado de la misma.
Otros dos nombres de significación menos positiva pero muy empleados son proximal y distal; el primero denota, en términos generales, la porción de una estructura más cerca del centro del cuerpo o de algún sitio importante de referencia; el segundo nombre se aplica a una porción más alejada. Estos términos son fáciles de aplicar a las extremidades y la cola. Dentro de la cabeza y el tronco, su uso es menos claro; sin embargo, cabe hablar de porciones proximal y distal de nervios (en relación con la médula o el encéfalo) y arterias (en relación con el corazón), etc.
Cada uno de estos adjetivos dan origen a los adverbios correspondientes, que significan movimiento en una dirección determinada: posteriormente, caudalmente.
Los nombres principales para denotar dirección (anterior, posterior, dorsal y ventral) se aplican con absoluta claridad a casi todos los vertebrados conocidos. Sin embargo, el hombre es excepción, por tratarse de una forma aberrante que adoptó la posición erguida, de manera que pueden aplicarse otros nombres de posición o dirección.
Es lamentable que así ocurra en la nomenclatura más empleada en anatomía médica (fig. 3). La cabeza y la cola del ser humano erguido están arriba y abajo, y no adelante y atrás, y se llaman superior e inferior, y no anterior y posterior (podría decirse craneal y caudal, pero los médicos odian estos términos tanto o más que quienes estudian anatomía comparada). Sin embargo, lo más grave es que se han utilizado mucho los nombres anterior y posterior en el ser humano y sin necesidad en lugar de dorsal y ventral, de manera que la porción de la espalda del ser humano suele llamarse posterior y la superficie del vientre anterior. Así, pues, estos dos adjetivos pueden tener significación contradictoria en anatomía humana especial y en sus acepciones más normales, y causar confusión innecesaria. Por ejemplo: cada nervio raquídeo tiene dos raíces (véase fig. 364). En la sala de disecciones las raíces en el cadáver humano suelen llamarse posterior y anterior. Sin embargo, si un neurólogo trabaja con ratas (por ejemplo) y trata de emplear la misma nomenclatura, caerá en un absurdo; una raíz no es más anterior o posterior que la otra, Sin embargo, en la rata y el hombre designar las raíces nerviosas con los adjetivos dorsal y ventral es razonable y lógico. Las costumbres consagradas por el tiempo, por más ilógicas que sean, son difíciles de vencer; sin embargo, en una revisión recientemente adoptada de nomenclatura anatómica humana se aceptó utilizar los nombres dorsal y ventral en estas circunstancias.
CONCEPTO DE HOMOLOGIA Incluso en los principios de la investigación zoológica se advirtió que en cada grupo principal de animales había una norma básica común del plano anatómico de la economía. En muchos miembros de un grupo, o en todos ellos, podían identificarse los mismos órganos, aunque a menudo muy modificados en volumen, forma, incluso función, en relación con el modo de vida. Al aceptar los biólogos el principio de la evolución, en la segunda mitad del siglo pasado, adquirió importancia verdadera el concepto de homología, o sea, la hipótesis de que órganos específicos de miembros vivientes de un grupo de animales proceden, aunque con modificaciones más o menos notables, de órganos básicamente idénticos presentes en el antecesor común. Durante decenios investigar homologías fue el propósito principal de la investigación zoológica.
Muchos resultados de estos estudios fueron nuevos e interesantes. Por ejemplo: se descubrió que los tres huesecillos auditivos del oído medio del ser humano (cap. 15) en edades más tempranas formaban parte del aparato mandibular de los peces, nuestros antecesores, y más temprano aún parecen haber sido parte del sostén de las branquias de vertebrados ancestrales. Los músculos con los cuales sonreímos o fruncimos el entrecejo provienen de los que un día ayudaron a los peces a bombear agua por las branquias.
Órganos homólogos son aquellos idénticos en la serie de formas que se estudian. Sin embargo, cabe preguntarse lo que significa idéntico. Sin razonar, tiende a suponerse que la misma masa verdadera de material, el miembro mismo, el pulmón o el hueso han pasado de unos a otros, generación tras generación, como bienes heredados. Esto es absurdo, pero este concepto ha influenciado notablemente, aunque en forma inconsciente, a muchos investigadores. En realidad, claro está, cada órgano vuelve a ser creado en cada generación, y la identidad entre homólogos se funda en la identidad o semejanza de los fenómenos evolutivos que los producen.
Los adelantos de la genética han brindado una base firme para interpretar estos fenómenos. Dependen de unidades hereditarias, los genes. Estas diminutas estructuras se cuentan por millares, cuando menos, en todas las células animales. El desarrollo de un animal dado es regido por los genes que recibe el huevo de los progenitores. Cada gen puede afectar el desarrollo de cierto número de estructuras o partes del cuerpo; a la inversa, cada órgano es influido en su desarrollo por muchos genes. Si los genes permanecen sin modificación de una generación a otra, el órgano producido será idéntico (aparte de efectos ambientales pasajeros en un individuo dado), y la homología es absoluta.
Sin embargo, en los genes ocurren cambios que se llaman mutaciones; modifican las estructuras que son originadas por los genes. Si las mutaciones producen efectos de escasa magnitud y ocurren sólo en algunos genes, el órgano se modificará poco, y la homología con el tipo original seguirá siendo patente. Sin embargo, si las mutaciones son muchas y de efecto notable, el órgano puede modificarse de manera radical y la ascendencia ser mucho menos manifiesta. En cierto sentido, el estudio de la homología de órganos es, sencillamente, estudiar los fenómenos producidos por los genes. Sí se conociera cabalmente la constitución genética de todos los tipos animales, para precisar la homología entre las estructuras podríamos fundamos en el grado de identidad de los genes que participan en su producción. Sin embargo, este no es asunto de importancia práctica, pues son poco probable que el campo de nuestros conocimientos se amplíe en la medida necesaria para lograr lo anterior,
Cabe preguntarse cuáles son los principales requisitos para establecer la homología. La función no es orientación fidedigna, pues órganos manifiestamente homólogos en los animales pueden tener funciones muy diferentes. La observación revela que la forma, el volumen o el color de una estructura brindan pocos datos positivos de identidad. La semejanza de posición anatómica enera1 y relaciones con órganos adyacentes es más útil para la identificación. Lo óptimo es la semejanza en la historia embriológica. Los fenómenos embriológicos en los vertebrados tienden a ser conservadores, y órganos que difieren mucho en el adulto pueden manifestar su homología a través de su semejanza en etapas en el embrión.
La homología se aplica, en general, a identidad estructural o anatómica. Algunos autores han propuesto que el concepto se amplíe para incluir la identidad funcional, Sin embargo, esto no se ha aceptado en general.
La palabra analogía denota muchas veces el equivalente funcional de la homología; órganos análogos los que poseen función semejante. Sin embargo, este término tiene limitaciones, pues en su sentido habitual significa que lOS órganos no son homólogos.
Por ejemplo, los pulmones y las branquias son análogos, pues se utilizan para la respiración, pero son estructuras por completo distintas.
ADAPTACION Y EVOLUCION Las modificaciones varias que han experimentado las estructuras de los vertebrados, y las diversas funciones que han asumido, resultan, claro está, de la evolución. No puede hacerse un estudio comparativo de los vertebrados sin formular algún concepto general sobre el carácter de los fenómenos de la evolución. La mayor parte de los cambios anatómicos y funcionales en el cuerpo de los vertebrados constituyen, en forma bastante clara, modificaciones adaptativas a diversos ambientes y modos de vida. Cabe preguntarse cómo se han producido estas adaptaciones. La explicación adecuada necesitaría una obra especial; sólo podemos indicar el carácter general de los problemas de base y la opinión actual más aceptada acerca de su interpretación.
A veces hablamos, sin pensarlo, de cambios adaptativos como si el animal los hubiera deseado o sus necesidades o deseos, por sí mismos, hubieran dado origen a nuevas estructuras o cambios estructurales. Pondremos un ejemplo: sería ventajoso para un pez poder caminar en la tierra, de manera que algunos peces crearon piernas; hubiera sido magnífico que los primeros antecesores de la vaca tuviesen dientes adecuados para masticar cereales y pasto, de manera que los dientes adquirieron rápidamente la estructura necesaria.
Es evidente que estas nociones son absurdas, Sin embargo, no dejan de tener relación con algunas teorías de la evolución que han gozado de aceptación, y siguen gozando de ella. Suponen que la evolución es un fenómeno anómalo; que los cambios han resultado de algún impulso interno del organismo, o del plan de alguna fuerza sobrenatural. Considerando que estas teorías no son científicas, no pueden refutarse científicamente. Sin embargo, tenemos derecho de buscar explicaciones más lógicas de la evolución, fundadas en hechos comprobados. Si alguien me dice que el funcionamiento del motor de mi automóvil es regulado por un demonio invisible que reside en él, no puedo probar que está equivocado. Sin embargo, nada gano añadiendo este demonio hipotético, y preferiría tratar de explicar el trabajo del motor en términos de principios mecánicos conocidos, por corrientes eléctricas y estructura explosiva de las moléculas de hidrocarburos.
Un intento más plausible para interpretar los cambios estructurales evolutivos fue el que presentó Lamarck hace más de un siglo; se funda en el concepto de que se heredan caracteres adquiridos por el efecto del uso y la falta de uso. Según esta hipótesis, cuando los antecesores de la jirafa estiraron el cuello para comer las hojas de las ramas más altas, el efecto del estiramiento se transmitió a los descendientes, generación tras generación, y el cuello alargado surgió de manera gradual en la norma hereditaria. Cuando los lagartos antecesores de la serpiente dejaron de usar las extremidades para la locomoción, el resultado acumulativo de la falta de uso fue la pérdida definitiva de las extremidades. Esta teoría interesante parece sencilla, lógica y natural. Sin embargo, las pruebas en que se apoya son escasas. En resumen, podemos decir que nadie, a pesar de repetidos intentos, ha podido encontrar pruebas valederas de un solo caso de transmisión de un carácter adquirido. Las estructuras útiles para un animal pueden aumentar en volumen y complejidad, y a menudo lo hacen, con el curso del tiempo, y las estructuras inútiles o poco usadas pueden disminuir. Sin embargo, no hay la menor prueba de que el uso o la falta de uso de alguna parte por un animal dado tenga efecto alguno en la constitución de los descendientes.
En los decenios últimos, la genética ha comprobado que los cambios evolutivos dependen de mutaciones; éstas pueden producir efectos de cierta magnitud, pero la mayor parte causan modificaciones pequeñas. Por ejemplo, una mutación en la mosca de la fruta quizá no tenga más efecto manifiesto que el desdoblamiento de una cerda. No se conocen a fondo las causas de las mutaciones, aunque en la actualidad estamos adquiriendo cierta noción de la estructura química de los materiales que intervienen, y de las influencias químicas y físicas (radiación, por ejemplo) que tienen papel importante en su producción.
Sin embargo, en lo que se refiere a las teorías de la evolución se destacan claramente dos hechos:
1) no hay pruebas de un plan o dirección en las mutaciones; parecen ocurrir bastante al azar, y no seguir una dirección dada; algunas posiblemente sean ventajosas; sin embargo, la mayor parte son claramente perjudiciales, a menudo mortales.
2) No hay pruebas de que las mutaciones guarden relación con el uso o la falta de uso de los órganos del cuerpo; los caracteres adquiridos por un animal no influyen sobre las mutaciones de los genes en las células sexuales, mutaciones cuyo efecto se transmite a los descendientes.
Así, pues, los fenómenos de mutación parecen consistir en cambios ciegos y al azar. Sin embargo, es evidente que la evolución de los vertebrados ha producido cambios útiles y adaptativos. Cabe preguntarse la forma en que estos cambios han sido producidos por la mutación.
Este problema fue resuelto por Darwin, en parte por lo menos, hace casi un siglo.
Claro está, desconocía los datos que hoy nos ha brindado la genética; sin embargo, supuso lógicamente que existían algunos mecanismos hereditarios, como los que hoy conocemos. Dada una serie de mutaciones al azar, la selección natural actuaría de manera preponderante para eliminar los tipos inadecuados y conservar las formas más adecuadas, en las cuales hubiese ocurrido en el plasma germinativo una mutación útil, o un grupo de ellas. Incluso los animales que menos se reproducen originan más individuos de los que pueden sobrevivir. Muchos están condenados a morir antes de convertirse en adultos y reproducirse, acto que es la norma estándar de éxito de la naturaleza. Qué individuos serán eliminados depende, en parte, del azar. Sin embargo, la observación y los experimentos indican que incluso las mutaciones pequeñas en dirección adaptativa poseen valor neto para la supervivencia, y muy pronto pueden tornarse dominantes en una especie. Esta selección natural de las mutaciones al azar que posean valor adaptativo parece ser un mecanismo fundamental de los cambios evolutivos.
Sin embargo, la evolución es un fenómeno mucho más complejo que una simple selección adaptativa de caracteres aparecidos en cierto individuo. Por ejemplo, puede señalarse que, en términos generales, llegan dos genes de cada tipo a las células del animal (un gen proviene de cada progenitor); si los miembros de un par de genes difieren en potencialidades, uno tiende a dominar al otro en las estructuras o funciones que regula. Es evidente que la selección no puede influir sobre el componente más débil de este par de genes, que recibe el nombre técnico de recesivo, a menos que por azar ambos componentes del par tengan el mismo carácter recesivo. Con pensar un poco se advierte de inmediato que, a causa de esta situación, es prácticamente imposible eliminar por completo un mutante recesivo de un grupo de animales cuando ya está establecido, aunque sea muy perjudicial. Es lógico suponer que cuando hay muchas variables de esta índole en una cepa, podrán presentarse circunstancias (sobre todo al modificarse el ambiente) en las cuales algunas variantes reprimidas o combinaciones de las mismas resulten por último muy ventajosas para el individuo en el cual se manifiesten, lo que se traducirá por un cambio evolutivo en la población global.
RELACIONES ENTRE SUPERFICIE Y VOLUMEN A menudo se advierte que en un grupo de animales las formas grandes y pequeñas difieren notablemente en el volumen relativo de diversos órganos o partes, El motivo de muchas de estas diferencias de proporción está en un principio geométrico tan obvio que a menudo pasa inadvertido: al modificarse el volumen de un animal (o de cualquier otro objeto), la superficie aumenta (o disminuye) proporcionalmente al cuadrado de las dimensiones lineales, en tanto que los volúmenes se modifican proporcionalmente al cubo de las dimensiones lineales.
Este principio tiene amplias aplicaciones, pues se descubren relaciones de superficie y volumen en muy diversos caracteres estructurales y funcionales de los vertebrados.
Citaremos ejemplos inmediatos:
1) la fuerza de la pata (al igual que toda columna de sostén) es función directa de la superficie transversal de sección, que varía como el cuadrado de las dimensiones lineales; en cambio, el peso que soporta es proporcional al cubo de las dimensiones lineales; en consecuencia, el elefante no puede tener patas como la gacela.
2) La cantidad de alimento que necesita un animal activo es proporcional, en números redondos, a su volumen; la cantidad de alimentos que el intestino puede absorber depende de la superficie del revestimiento intestinal; en consecuencia, en los animales voluminosos el intestino es muy largo o de estructura complicada, y brinda mayor superficie interna para la digestión.
NOMENCLATURA Quien estudia morfología de los vertebrados se enfrenta a un conjunto atemorizador de nombres raros de estructuras anatómicas. Esto es lamentable, pero inevitable. Los vertebrados poseen muchos órganos, y para gran parte de ellos no hay un nombre de uso diario. Incluso cuando lo haya, a menudo es vago y no tiene significación precisa en el lenguaje corriente. Además, conviene tener algún sistema internacional de nomenclatura que entiendan en el mismo sentido los científicos de todos los países.
Cuando comenzó a estudiarse anatomía, lo normal era que todos los trabajos eruditos se escribieran en latín. En consecuencia, los nombres latinos entonces en uso se aplicaban a estructuras anatómicas; si no había un nombre útil, se creaba uno con raíces griegas o latinas y se latinizaba. En el apéndice II se hacen algunas observaciones acerca de la formación de nombres anatómicos. En la actualidad, el latín ha dejado de ser lenguaje internacional en lo que se refiere a las obras científicas generales de texto. Sin embargo, siguen empleándose nombres anatómicos en latín. No podemos eliminarlos, aunque a menudo los empleamos en forma algo castellanizada; por ejemplo: hablamos del músculo deltoides del hombro y no del musculus deltoideus, o del hueso parietal y no del os parietalis
El latín es una lengua que posee declinaciones, y los nombres con distintas terminaciones se expresan no sólo singular y plural, sino también diversos casos y un sistema bastante arbitrario de géneros. Hasta hace poco, todo estudiante de colegios superiores sabía algo de gramática latina, y el manejo de los nombres latinos no entrañaba dificultad. Pero ya no es así, y ello es lamentable. Por fortuna, el número en terminaciones para nombres y adjetivos de uso corriente en la nomenclatura anatómica es limitado, y puede aprenderse fácilmente.
Es aceptado en nomenclatura anatómica que cuando un órgano existe en los mamíferos sobre todo en el hombre debe darse este nombre a la misma estructura en otros animales. Por ejemplo: el hombre y muchos mamíferos poseen clavícula, y el elemento equivalente en la estructura del hombro debe llevar el mismo nombre en reptiles, anfibios o peces, aunque su aspecto sea por completo distinto. Sin embargo, en ocasiones se hace una identificación demasiado apresurada y se aplica equivocadamente un nombre. Los peces teleósteos poseen un hueso de situación semejante a la clavícula, y se acostumbraba llamarlo clavícula; sin embargo, hoy sabemos que los teleósteos perdieron la clavícula verdadera, y el hueso de este sitio es diferente (el cleitro). Si la homología es discutible, conviene dar un nombre distinto a la estructura que se trata. Por ejemplo: en el muslo de los reptiles hay un músculo que pudiera ser homólogo del sartorio de los mamíferos; sin embargo, como hay dudas acerca de la homología, el músculo de los reptiles suele llamarse músculo ambiens. Claro está, si una estructura observada en un grupo inferior no tiene equivalente en los mamíferos, debe idearse un nombre adecuado para ella.
Aunque la nomenclatura anatómica ha sido, en general, un sistema bastante estable y uniforme, es lógico que hayan surgido diferencias entre diversas escuelas de trabajo y en distintos países. Movida por el deseo plausible de lograr uniformidad, la Asociación Alemana de Anatomistas, reunida en un congreso en Basilea hace algunos decenios, produjo un sistema amplio de nombres con la esperanza de que se adoptaran mundialmente para la anatomía humana.
Esta nomenclatura, que suele llamarse
Norma Anatómica de Basilea (BNA) fue adoptada por las escuelas de medicina y se ha empleado mucho en los trabajos médicos, Por desgracia, algunos nombres se eligieron inadecuadamente; ya hemos visto las dificultades que surgieron para los nombres que denotaban posición corporal. Un congreso internacional de anatomistas reunido en París en 1955 creó una nueva nomenclatura, mejorando bastante la anterior; en la actualidad, las revisiones incluidas en la Nómina Anatómica de París (PNA) están substituyendo gradualmente a los nombres antiguos en las escuelas de medicina.
____________________________________________________________