Fuente: FisicaNet
http://www.fisicanet.com.ar/
_________________________________________________________
Apunte de Metabolismo:
Moneda energética, energía liberada en reacciones exergónicas muchas veces acopladas.
ATP
En las células todas las actividades biosintéticas, muchos de sus procesos de transporte y una diversidad de otras actividades requieren energía. Una gran proporción de esta energía es suministrada por una sola sustancia: el adenosín trifosfato o ATP. La glucosa y otros carbohidratos son formas de almacenamiento de energía y también formas en las que se transfiere energía de célula a célula y de organismo a organismo. Son las reservas de energía, el ATP es la moneda energética de la célula que puede gastarse de inmediato.
La molécula de ATP está formada por la base nitrogenada adenina, el azúcar de cinco carbonos ribosa y tres grupos fosfato. Los tres grupos fosfato están unidos por dos enlaces covalentes que se rompen con facilidad, produciendo cada uno aproximadamente 7 kilocalorías de energía por mol.
En los sistemas biológicos, las reacciones endergónicas (requieren energía), como las de biosíntesis, se producen gracias a la energía liberada en las reacciones exergónicas (liberan energía) con las que están acopladas . En la mayoría de las reacciones acopladas, el ATP es el intermediario que conduce la energía de una reacción a otra.
La oxidación de la glucosa es una fuente principal de energía en la mayoría de las células. Cuando la glucosa se degrada en una serie de pequeños pasos por medio de enzimas, una proporción significativa de la energía contenida en la molécula vuelve a empaquetarse en los enlaces fosfato de las moléculas de ATP.
La primera fase en la degradación de la glucosa es la glucólisis que se efectúa en el citoplasma de la célula. La segunda fase es la respiración aeróbica, que requiere oxígeno y, en las células eucarióticas, tiene lugar en las mitocondrias. La respiración comprende el ciclo de Krebs y el transporte en cadena de electrones acoplado al proceso de fosforilación oxidativa. Todos estos procesos están íntimamente relacionados.
En condiciones anaeróbicas, el proceso de fermentación transforma al ácido pirúvico producido por la glucólisis o en etanol o en ácido láctico, dependiendo del organismo y el medio.
Es posible saber cómo y en qué cantidad la energía química, originalmente presente en la molécula de glucosa, se recupera en forma de ATP en el curso de la degradación de la molécula de glucosa. Así, es posible calcular el rendimiento energético global de la oxidación de la glucosa, que puede dar como resultado un máximo de 38 moléculas de ATP. La actividad de la glucólisis y la respiración están reguladas de acuerdo con las necesidades energéticas de la célula
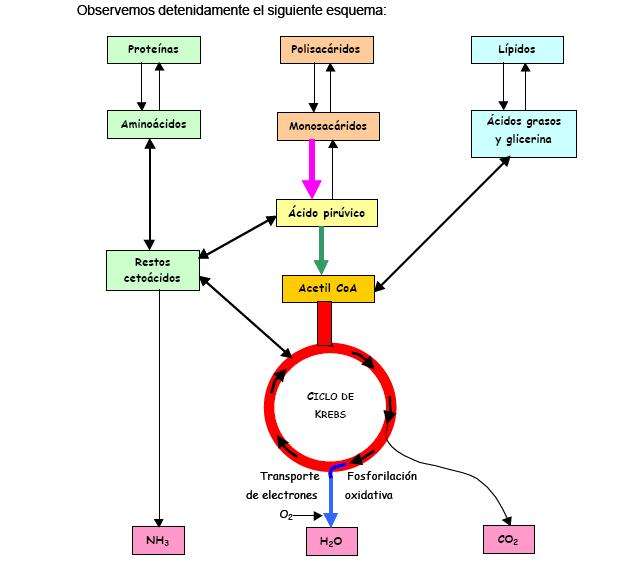
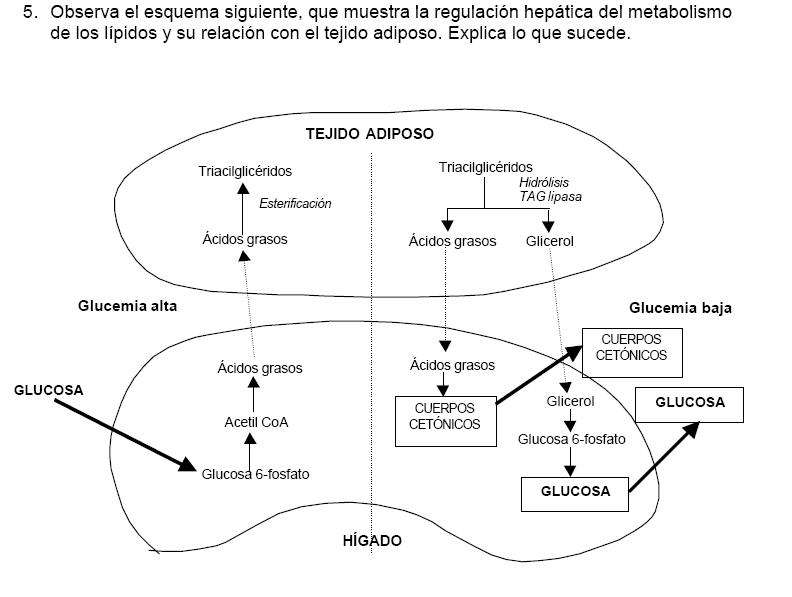
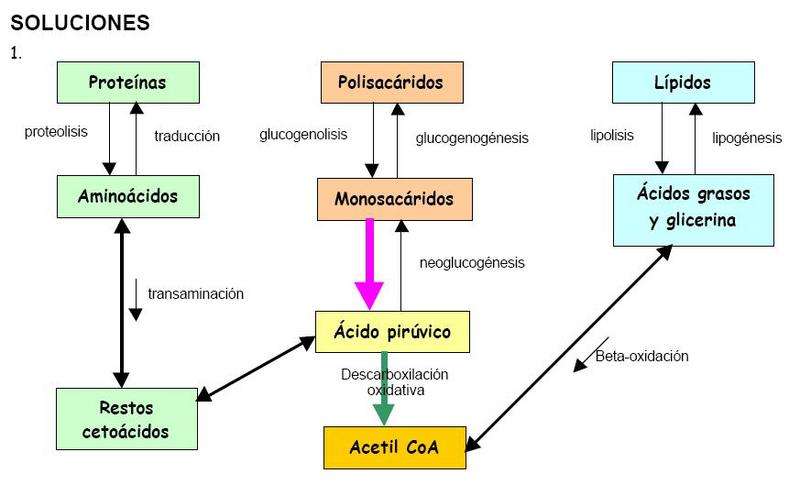
Otras moléculas alimenticias, que incluyen a las grasas, los polisacáridos y las proteínas, pueden ser también degradadas a compuestos que pueden ingresar en las vías centrales -glucólisis y ciclo de Krebs- en diferentes pasos. La biosíntesis de compuestos orgánicos utiliza los compuestos precursores derivados de intermediarios en la secuencia respiratoria y es impulsada por la energía derivada de esos procesos. Así, otras vías catabólicas y anabólicas están íntimamente interrelacionadas.
Autor: Diana Victoria Netto.
Bibliografía:
Apuntes de clase - Facultad de Agronomía - U.B.A..
A. Lehninger, D, Nelson y M. Cox. "Principios de bioquímica". Editorial Omega, 2000.
Helena Curtis - N. Sue Barnes. "Biología". Sexta edición en español.
____________________________________________________