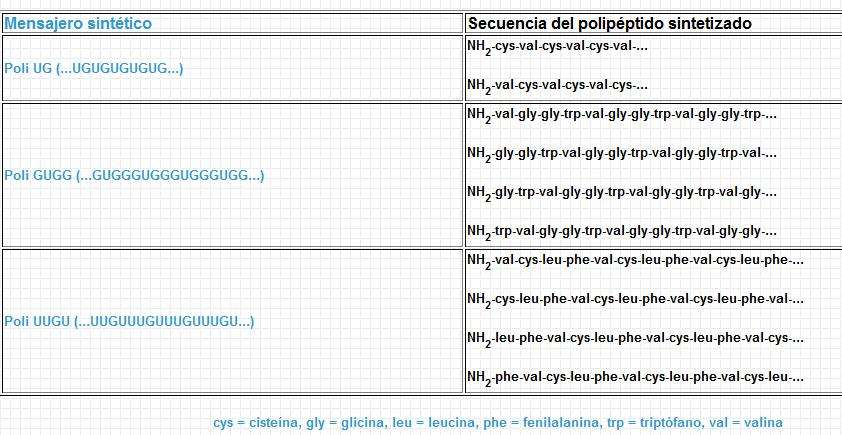
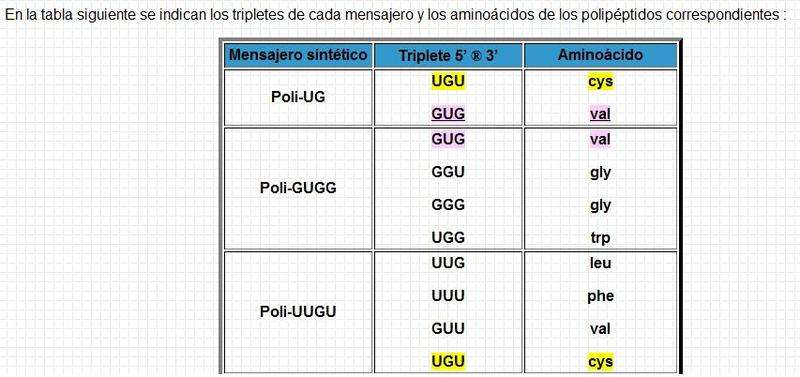
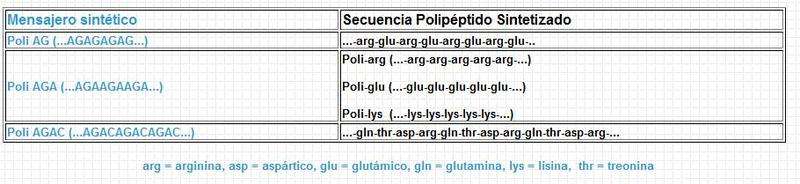
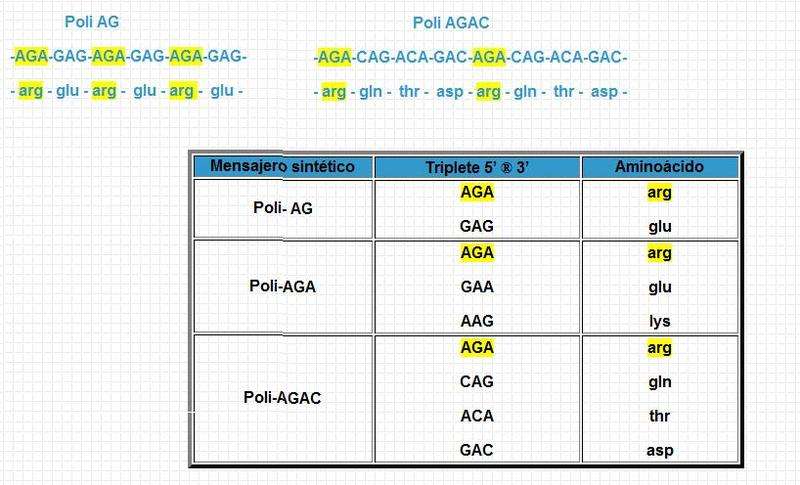
__________________________________________
3.7.- Cuando se suplementa con los compuestos A, B, C, D, E, F, G, H e I el medio mínimo en el que se cultivan las cepas mutantes nutricionales de Neurospora 1 al 9 se obtienen los siguientes resultados:

_______________________________________________________