Capítulo 12.
En el jardín de un monasterio: el comienzo de la genética
Las nociones más tempranas
acerca de la herencia biológica giraban alrededor de la inquietud de
conocer cómo se transmiten las características hereditarias de
generación en generación.
La revolución en la genética
se produjo cuando el concepto de mezcla fue reemplazado por el concepto de factor
o unidad de la herencia. La gran contribución de Mendel fue demostrar
que las características heredadas son llevadas en unidades discretas
que se reparten por separado -se redistribuyen- en cada generación. Estas
unidades discretas, que Mendel llamó elemente, son los que hoy conocemos
como genes.
La hipótesis de que cada individuo lleva un par de factores para cada
característica y que los miembros del par segregan -es decir, se separan-
durante la formación de los gametos, se conoce como primera ley de Mendel,
o principio de segregación. La segunda ley de Mendel, o principio de
la distribución independiente, establece que, cuando se forman los gametos,
los alelos del gen para una característica dada segregan independientemente
de los alelos del gen para otra característica.
Las mutaciones son cambios abruptos
en el genotipo. Son la fuente primaria de las variantes genéticas estudiadas
por Mendel. Diferentes mutaciones en un gen único incrementan la diversidad
de alelos de ese gen en la población. En consecuencia, la mutación
aporta la variabilidad existente entre los organismos, que es la materia prima
para la evolución.
Las investigaciones realizadas por Mendel no fueron valoradas por sus colegas
científicos y tuvieron que esperar mucho tiempo hasta ser "redescubiertas".
Las décadas que siguieron al redescubrimiento de los trabajos de Mendel
fueron muy ricas en estudios genéticos que resultarían de enorme
importancia.
Concepciones acerca de la herencia
Transcurrieron muchos siglos
en los que diferentes creencias y mitos predominaron sobre las explicaciones
científicas. A mediados del siglo XIX, ya se sabía que los óvulos
y los espermatozoides son células especializadas y que, tanto el óvulo
como el espermatozoide, contribuyen a las características hereditarias
del nuevo individuo. Pero ¿cómo, estas células especiales
llamadas gametos, son capaces de transmitir las centenas de características
involucradas en la herencia? La herencia mezcladora, que sostenía que
las características de los progenitores se mezclaban en la progenie,
como en una mezcla de dos fluidos, fue una de las hipótesis. Sin embargo,
esta explicación no tenía en cuenta la persistente herencia de
ciertas variantes que indudablemente ocurría.
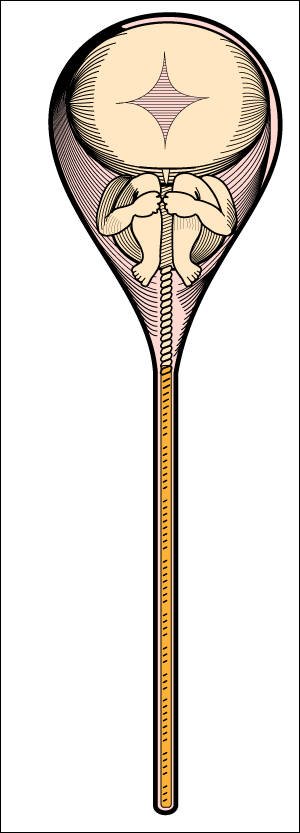
Este es un homúnculo ("hombrecito"), futuro ser humano en miniatura,
dentro de un espermatozoide.
Imagen de lo que creían ver los animalculistas o espermistas de los siglos XVII y XVIII cuando miraban espermatozoides
Los principios de Mendel
La primera ley de Mendel, o
principio de segregación establece que cada individuo lleva un par de
factores para cada característica y que los miembros del par segregan
-es decir, se separan- durante la formación de los gametos.
Si los miembros del par son
iguales, se dice que el individuo es homocigota para la característica
determinada por ese gen; si son diferentes, el individuo es heterocigota para
esa característica. Las diferentes formas de un mismo gen son conocidas
como alelos.
La constitución genética
de un organismo se denomina genotipo. Sus características externas observables
se conocen como fenotipo. Un alelo que se expresa en el fenotipo de un individuo
heterocigota, con exclusión del otro alelo, es un alelo dominante; aquel
cuyos efectos no se observan en el fenotipo del heterocigota es un alelo recesivo.
En los cruzamientos que involucran a dos individuos heterocigotas para el mismo
gen, la relación en la progenie del fenotipo dominante con respecto al
recesivo es 3:1.
Mendel cruzó una planta de guisante pura de semillas amarillas con una planta pura de semillas verdes, transfiriendo el polen de las anteras de las flores de una planta a los estigmas de las flores de otra planta. Estas plantas constituyeron la generación progenitora (P). Las flores así polinizadas originaron vainas de guisantes que contenían solamente semillas amarillas. Estos guisantes -que son semillas- constituyeron la generación F1. Cuando las plantas de la F1 florecieron, las dejó autopolinizarse. Las vainas que se originaron de las flores autopolinizadas (generación F2) contenían tanto semillas amarillas como verdes, en una relación aproximada de 3:1, o sea aproximadamente 3/4 eran amarillas y 1/4 verdes.
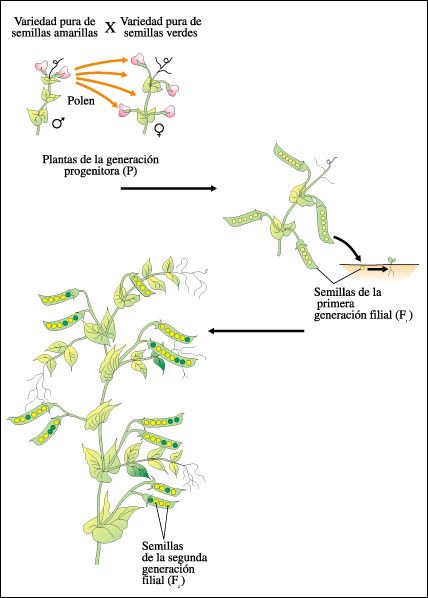
Esquema de los experimentos de Mendel.
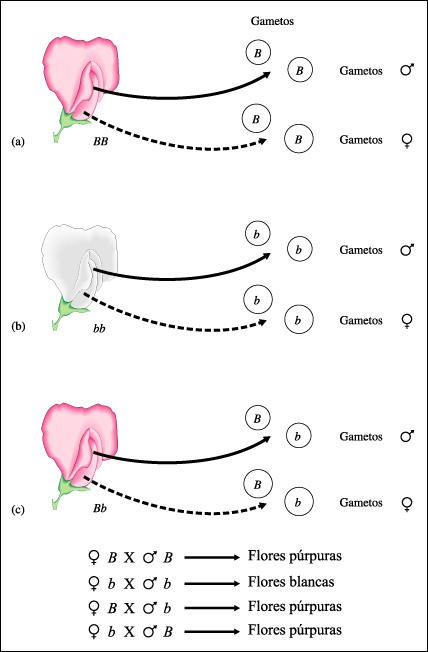
Esquema de la segregación de los alelos durante la formación de los gametos.
Una planta de guisante homocigota
para flores púrpuras, se representa como BB en símbolos genéticos
ya que el alelo para flor púrpura es dominante (B). Esta planta BB, sólo
produce gametos, ya sean femeninos o masculinos, con el alelo para flor púrpura
(B). Del mismo modo, una planta de guisante de flores blancas es homocigota
recesiva (bb) y solamente produce gametos femeninos o masculinos con el alelo
para flor blanca (b). Finalmente, una planta heterocigota (Bb) posee flores
púrpura ya que el alelo para flor púrpura (B) es dominante sobre
el alelo para flor blanca (b); esta planta produce la mitad de los gametos con
el alelo B y la otra mitad, con el alelo (b), ya sea que se trate de gametos
femeninos o masculinos.
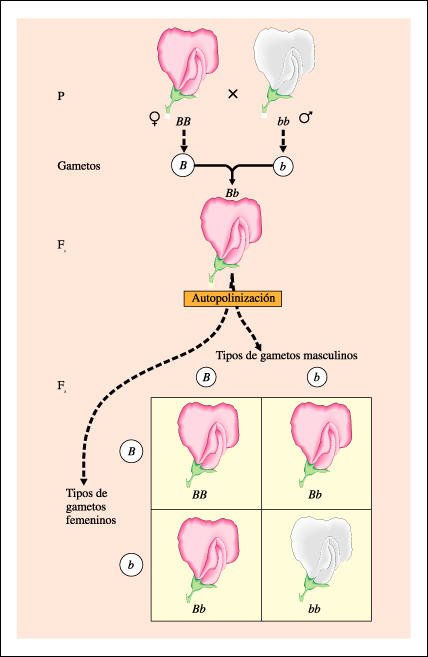
Esquema del principio de segregación de Mendel.
Se muestran las generaciones
F1 y F2 después de un cruzamiento entre plantas de la generación
P: una planta de guisante homocigota dominante para flores púrpuras (BB)
y una planta homocigota recesiva para flores blancas (bb).
El fenotipo de la progenie de
este cruzamiento-la generación F1- es púrpura, pero su genotipo
es Bb. La F1 heterocigota produce cuatro tipos de gametos: masculinos B, femeninos
B, masculinos b y femeninos b, en proporciones iguales. Cuando esta planta se
autopoliniza, los gametos masculinos y los femeninos, B y b, se combinan al
azar y forman, en promedio 1/4 BB (púrpura), 2/4 (o 1/2) Bb (púrpura)
y 1/4 bb (blanco). La relación genotípica subyacente 1:2:1 es
la que da cuenta de la relación fenotípica de tres dominantes
(púrpura) a un recesivo (blanco), que se expresa como 3:1. La distribución
de las variantes en la F2 se muestra en un tablero de Punnett, que recibió
su nombre del genetista inglés que utilizó este tipo de diagrama
para el análisis de las características determinadas genéticamente.
Un cruzamiento de prueba, en el cual un individuo con una característica fenotípica dominante -pero con un genotipo desconocido- se cruza con un individuo homocigota para el alelo recesivo, revela el genotipo desconocido. Si en un cruzamiento de prueba que involucra a un gen aparecen en la progenie los dos fenotipos posibles, el individuo probado es heterocigota; si, en cambio, en la progenie solamente aparece el fenotipo dominante, el individuo es homocigota para el alelo dominante.
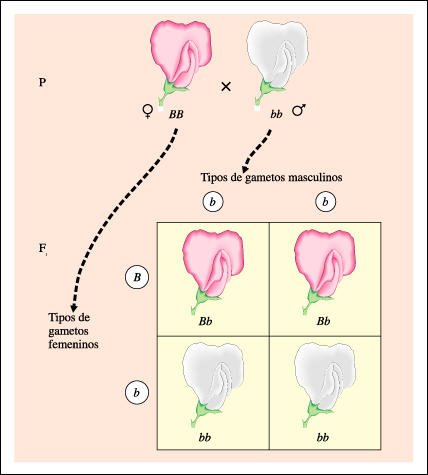
Cruzamiento de prueba.
Para que una flor de guisante
sea blanca, la planta debe ser homocigota para el alelo recesivo (bb). Pero
una flor de guisante púrpura puede ser producida por una planta de genotipo
Bb o por una de genotipo BB. ¿Cómo se podría distinguir
una de otra? Los genetistas resuelven este problema cruzando estas plantas con
otras que sean homocigotas recesivas. Este tipo de experimento se conoce como
cruzamiento de prueba. Como se muestra aquí, la relación fenotípica
en la generación F1 -de igual número de plantas con flor púrpura
que de plantas con flor blanca (1:1)- indica que la planta con flor púrpura
utilizada como progenitor en el cruzamiento de prueba era heterocigota.
El segundo principio de Mendel, el principio de la distribución independiente, se aplica al comportamiento de dos o más genes diferentes. Este principio establece que los alelos de un gen segregan independientemente de los alelos de otro gen. Cuando se cruzan organismos heterocigotas para cada uno de dos genes que se distribuyen independientemente, la relación fenotípica esperada en la progenie es 9:3:3:1.
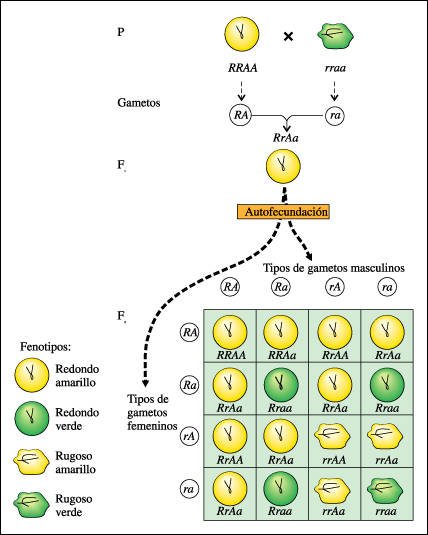
Esquema del principio de la distribución independiente de Mendel.
Una planta homocigota para semillas
redondas (RR) y amarillas (AA) se cruza con una planta que tiene semillas rugosas
(rr) y verdes (aa). Toda la generación Fl tiene semillas redondas y amarillas
(RrAa). Veamos en qué proporciones aparecen las variantes en la generación
F2. De las 16 combinaciones posibles en la progenie, 9 muestran las dos variantes
dominantes (RA, redonda y amarilla), 3 muestran una combinación de dominante
y recesivo (Ra, redonda y verde), 3 muestran la otra combinación (rA,
rugosa y amarilla) y 1 muestra las dos recesivas (ra, rugosa y verde). Esta
distribución 9:3:3:1 de fenotipos siempre es el resultado esperado de
un cruzamiento en que intervienen dos genes que se distribuyen independientemente,
cada uno con un alelo dominante y uno recesivo en cada uno de los progenitores.
Mutaciones
A partir de los trabajos de
Mendel se realizaron numerosas investigaciones sobre la herencia. El botánico
Hugo de Vries, en sus estudios sobre herencia mendeliana en la planta "hierba
del asno", también llamada "diego de noche", encontró
que la herencia en esta especie generalmente era ordenada y predecible, como
ocurría en el guisante. Sin embargo, ocasionalmente aparecía alguna
variante que no estaba presente ni en los progenitores ni en ningún antecesor
de esta planta.
De Vries conjeturó que
estas variantes surgían como resultado de cambios súbitos en los
genes y que la variante producida por un gen cambiado se transmitía luego
a la progenie, como lo hace cualquier otra característica hereditaria.
De Vries denominó mutaciones a estos cambios hereditarios repentinos,
y a los organismos que exhibían estos cambios, mutantes. Los conceptos
propuestos por de Vries resultaron erróneos, el concepto de mutación
como fuente de la variación genética demostró ser de suma
importancia, aunque la mayoría de sus ejemplos no eran válidos.
Hoy se sabe que las mutaciones
son cambios abruptos en el material genético. Como resultado de las mutaciones,
existe una amplia gama de variabilidad en las poblaciones naturales. En un ambiente
heterogéneo o cambiante, una variación determinada puede darle
a un individuo o a su progenie una ligera ventaja. En consecuencia, aunque las
mutaciones no determinan la dirección del cambio evolutivo, constituyen
la fuente primaria y constante de las variaciones hereditarias que hacen posible
la evolución.
La influencia de Mendel
Los trabajos de Mendel no fueron
interpretados en toda su dimensión cuando fueron presentados a la comunidad
científica. No fue hasta 1900 que los biólogos aceptaron los hallazgos
de Mendel. En un solo año, su trabajo fue redescubierto por tres científicos
quienes, en forma independiente, habían hecho experimentos similares
y estaban revisando la literatura especializada para confirmar sus resultados.
Durante los 35 años en que el trabajo de Mendel permaneció en
la oscuridad se había efectuado un considerable progreso en la microscopía
y, en consecuencia, en el estudio de la estructura celular. Durante este período,
se descubrieron los cromosomas y se observaron y describieron por primera vez
sus movimientos durante la mitosis. Durante estos años, también
se descubrió el proceso por el cual se forman los gametos y los sucesos
de la meiosis fueron rápidamente relacionados con los principios mendelianos
de la herencia. En las décadas que siguieron al redescubrimiento del
trabajo de Mendel se realizó una enorme cantidad de estudios genéticos.


