Capítulo 13.
Extensión de la genética mendeliana
Los trabajos de Mendel fueron
redescubiertos en Europa en 1900 por Hugo de Vries y otros científicos
y atrajeron una gran atención en todo el mundo, estimulando muchos estudios
de investigadores que buscaban confirmar y extender sus observaciones. El redescubrimiento
de los trabajos de Mendel fue el catalizador de muchos nuevos descubrimientos
en genética que condujeron a la identificación de los cromosomas
como los portadores de la herencia. Sin embargo, algunas de las conclusiones
de Mendel debieron ser modificadas.
Si bien muchas de las características
se heredan de acuerdo con las leyes establecidas por Mendel, otras, tal vez
la mayoría, siguen patrones de herencia más complejos. Ciertas
interacciones entre los alelos, interacciones entre los genes, e interacciones
con el medio ambiente explican gran parte de estas desviaciones de los principios
mendelianos.
Muchas veces, en los cromosomas
ocurren cambios que, según afecten su número o estructura, se
clasifican como alteraciones cromosómicas numéricas o alteraciones
cromosómicas estructurales, respectivamente. A veces, estas alteraciones,
o mutaciones, tienen consecuencias perjudiciales para los individuos, pues alteran
su viabilidad o su fertilidad. Otras veces, sin embargo, los cambios cromosómicos
se mantienen como parte de la variabilidad genética entre los organismos
y contribuyen al cambio evolutivo y al origen de nuevas especies.
Genes y cromosomas
Un fuerte apoyo a la hipótesis
de que los genes están en los cromosomas, provino de los estudios hechos
por el genetista Morgan y su grupo en la mosquita de la fruta D. melanogaster.
Dado que es fácil de criar y mantener, la Drosophila ha sido usada en
una variedad de estudios genéticos. Esta mosca tiene 4 pares de cromosomas;
3 pares -los autosomas - son estructuralmente iguales en ambos sexos, pero el
cuarto par, los cromosomas sexuales, es diferente. En la mosquita de la fruta,
como en muchas otras especies (incluidos los humanos), los dos cromosomas sexuales
son XX en las hembras y XY en los machos.
En el momento de la meiosis,
los cromosomas sexuales, al igual que los autosomas, segregan. Cada óvulo
recibe un cromosoma X, pero la mitad de los espermatozoides recibe un cromosoma
X y la otra mitad, un cromosoma Y. Así, en Drosophila, en los humanos
y en muchos otros organismos (aunque no en todos), es el gameto paterno el que
determina el sexo de la progenie.
En los primeros años del siglo XX, los experimentos de cruzamientos de Drosophila mostraron que ciertas características están ligadas al sexo, o sea, que sus genes se encuentran en los cromosomas sexuales. Los genes ligados al X dan lugar a un patrón de herencia particular. En los machos, como no hay otro alelo presente, la existencia de un alelo recesivo en el cromosoma X es suficiente para que la característica se exprese en el fenotipo. Por oposición, una hembra heterocigota para una variante recesiva ligada al X portará esa variante, pero ésta no se manifestará en su fenotipo.
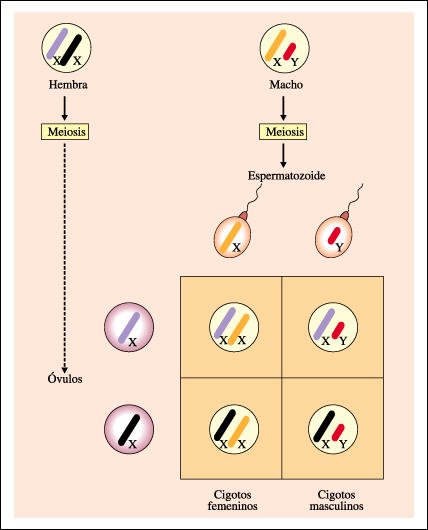
Determinación del sexo en organismos (como el ser humano y las moscas del género Drosophila) en los cuales el macho es heterogamético.
En la meiosis femenina, cada óvulo recibe un cromosoma X. En la meiosis masculina, cada espermatozoide puede recibir un cromosoma X o un cromosoma Y. Si un espermatozoide que lleva un cromosoma X fecunda al óvulo, el cigoto dará lugar a una hembra (XX); si un espermatozoide que lleva un cromosoma Y fecunda al óvulo, el cigoto dará lugar a un macho (XY).
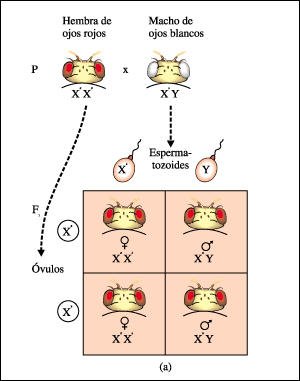
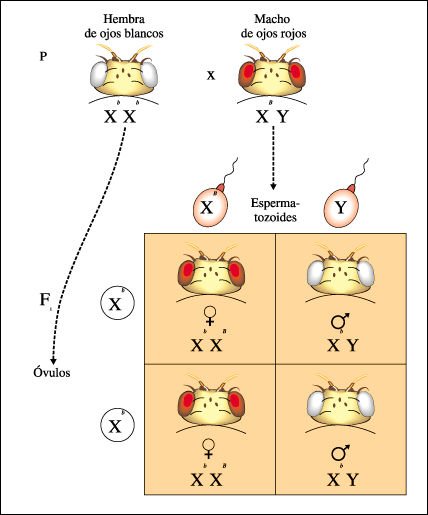
Diagramas del tablero de Punnett que representan los experimentos realizados después de descubrir un macho de Drosophila de ojos blancos.
Diagramas del tablero de Punnett
que representan los experimentos realizados después de descubrir un macho
de Drosophila de ojos blancos.
Morgan cruzó primero una hembra homocigota de ojos rojos con el macho
de ojos blancos; toda la progenie tuvo ojos rojos.
a) La característica
ojos blancos es menos común en las moscas y está representada
por una b, y B simboliza el alelo salvaje para ojos rojos. Como el gen está
localizado en el cromosoma X, los alelos se designan comúnmente con superíndices.
13-6b
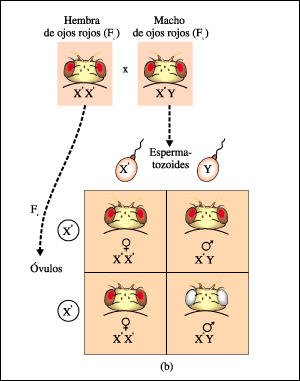
b) Morgan, luego apareó una hembra de ojos rojos de la Fl con un macho de ojos rojos de la Fl. Aunque en la generación F2 hubo machos de ojos rojos y de ojos blancos, todas las hembras F2 tuvieron ojos rojos, sugiriendo la existencia de una relación entre la herencia del color de los ojos y el comportamiento de los cromosomas sexuales.
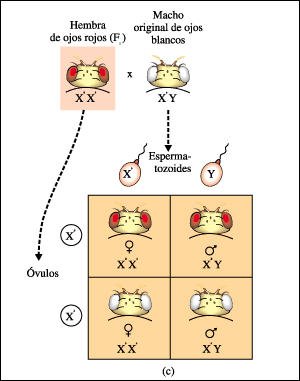
c) Un cruzamiento de prueba
entre una hembra Fl de ojos rojos y el macho original de ojos blancos produjo
mosquitas de ojos rojos y de ojos blancos en ambos sexos. Esto llevó
a la conclusión de que el gen para el color de ojos debía estar
localizado en el cromosoma X. El alelo para ojos rojos (B) es dominante y el
alelo para ojos blancos (b) es recesivo.
Por efecto del entrecruzamiento meiótico, los alelos se intercambian entre cromosomas homólogos. Estas recombinaciones ocurren porque: 1) los genes están dispuestos en un orden lineal fijo a lo largo de los cromosomas, y 2) los alelos de un gen dado están en sitios (loci) específicos en cromosomas homólogos. Los mapas cromosómicos, que muestran las posiciones relativas de los loci de los genes a lo largo de los cromosomas, han sido construidos a partir de datos de recombinación obtenidos de experimentos de cruzamientos. Aunque algunos genes se distribuyen independientemente, como predice la segunda ley de Mendel, otros tienden a permanecer juntos. Cuando los genes no se distribuyen independientemente, se dice que están ligados y su grado de ligamiento depende de la distancia relativa que existe entre ellos.
Progenie de un cruzamiento entre una mosquita de la fruta hembra de ojos blancos y un macho de ojos rojos, ilustrando lo que ocurre cuando el alelo recesivo está localizado en un cromosoma X.
Las hembras Fl, con un cromosoma X materno y otro paterno, son heterocigotas (Xb XB) y presentan ojos rojos. Pero los machos Fl, con su único cromosoma X recibido de la madre, llevan el alelo recesivo b, y serán todos de ojos blancos, dado que el cromosoma Y no lleva gen para color de ojos. Así, el alelo recesivo en el cromosoma X heredado de la madre se expresa en los machos de la progenie.
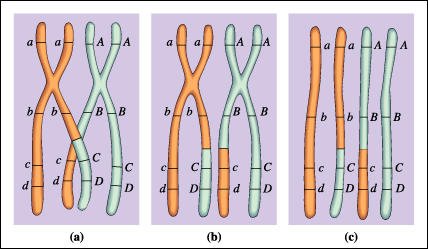
Entrecruzamiento
a) y b) El entrecruzamiento
se inicia cuando se aparean las cromátides homólogas, al inicio
de la meiosis I. Luego se produce la ruptura de las cromátides y los
extremos de cada una de ellas se unen con los de su homóloga. De esta
manera, los alelos se intercambian entre los cromosomas.
c) Como resultado de este proceso, los cromosomas homólogos tienen combinaciones
de alelos diferentes de las iniciales.
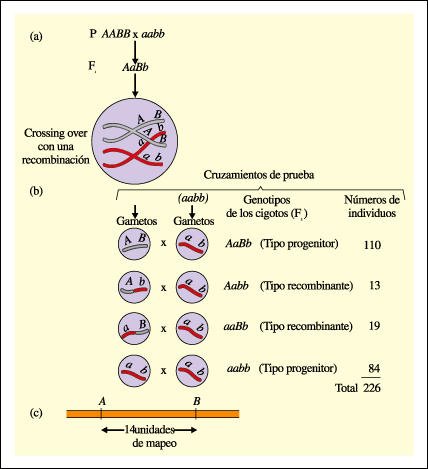
Determinación de la distancia de mapeo entre dos genes del mismo cromosoma.
a. Cuando un individuo homocigota
dominante para dos genes localizados en el mismo par de cromosomas homólogos
(AABB) se cruza con uno homocigota recesivo (aabb), la progenie F1 será
toda heterocigota para ambos genes (AaBb). Si hay entrecruzamiento durante la
meiosis, en el heterocigota los alelos de las cromátides de los dos homólogos
pueden intercambiarse y, como resultado de la recombinación, formarse
cuatro tipos diferentes de gametos. Los gametos progenitores -AB y ab- y los
gametos de tipo recombinante -Ab y aB-.
b. Apareamiento entre el heterocigota
de la generación F1 y un individuo homocigota recesivo (cruzamiento de
prueba).
c. La cantidad de recombinantes (13 + 19=32) dividida por la cantidad total de descendientes indica el porcentaje de recombinación (32 / 226 = 0,14), se define como la distancia de mapeo entre los genes. Entonces, los genes A y B están a una distancia de 14 unidades de mapeo.
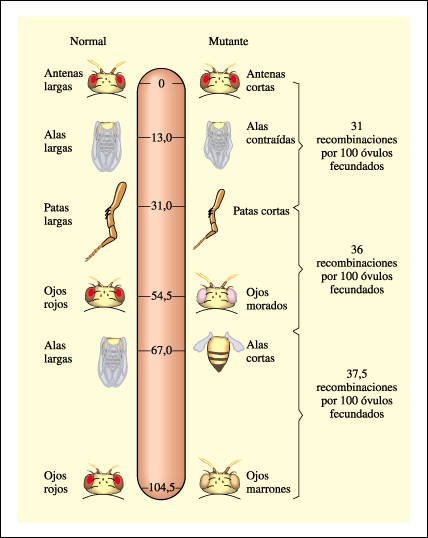
Porción de un mapa cromosómico de Drosophila melanogaster que muestra las posiciones relativas de algunos de los genes en el cromosoma 2, calculadas por la frecuencia de recombinaciones. Como puede verse, más de un gen puede afectar a una sola característica, como el color de los ojos.
Ampliando el concepto de gen
A medida que avanzaba la edad
de oro de la genética, lo nuevos estudios mostraban que los patrones
hereditarios no siempre son tan simples y directos. Si bien los principios mendelianos
constituyen la base para predecir los resultados de cruzamientos simples, las
excepciones, aunque no invalidan las leyes de Mendel, son abundantes. Ciertas
interacciones entre alelos explican gran parte de estas desviaciones de los
principios mendelianos. Aunque la interacción de la gran mayoría
de los alelos ocurre según la modalidad dominante-recesivo, en algunos
casos existe dominancia incompleta y codominancia. Además, aunque sólo
dos alelos están presentes en cualquier individuo diploide, en una población
de organismos un solo gen puede tener alelos múltiples, como resultado
de una serie de diferentes mutaciones de ese gen. La interacción entre
genes puede originar fenotipos nuevos y, en algunos casos, los genes pueden
presentar epístasis, es decir, uno de ellos modificar el efecto del otro.
Como resultado, se alteran las proporciones fenotípicas esperadas según
las leyes de Mendel. Asimismo, un solo gen puede afectar dos o más características
que aparentemente no están relacionadas; esta propiedad de un gen se
conoce como pleiotropía. En muchas características, la expresión
fenotípica está influida por varios genes; este fenómeno
se conoce como herencia poligénica. Los rasgos con este tipo de herencia
muestran variación continua y su estudio se realiza mediante curvas que
describen su distribución en las poblaciones.
Cuando la expresión de un gen se altera por factores del ambiente, o por otros genes, dos resultados son posibles. En primer lugar, el grado en que se expresa un genotipo particular en el fenotipo de un individuo puede variar. A este efecto se lo denomina expresividad variable. Frecuentemente, existe gran variabilidad en la expresividad de un gen entre los miembros de una misma familia. Además, la proporción de individuos que muestran el fenotipo correspondiente a un genotipo particular puede ser menor que la esperada: en este caso se dice que el genotipo muestra penetrancia incompleta.
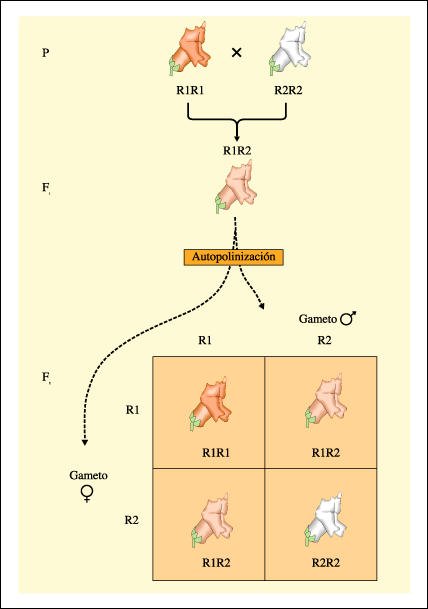
Un cruzamiento entre una planta boca de dragón con flores rojas (R1R1) y una con flores blancas (R2R2).
En este caso, ningún alelo es dominante. La flor del heterocigota presenta un fenotipo intermedio entre los dos colores.
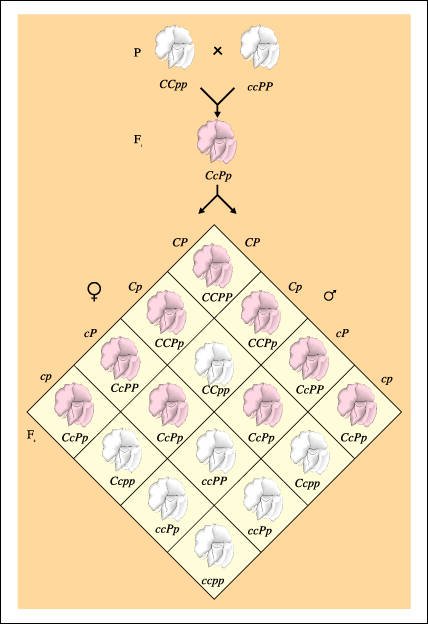
Epístasis en arvejillas de olor.
Cuando se cruzan dos variedades diferentes de arvejillas de olor de flores blancas, todas las plantas Fl tienen flores púrpuras. En la generación F2, la relación de plantas con flores púrpuras y flores blancas es 9:7. El color púrpura se debe a la presencia de ambos alelos dominantes, P y C; el homocigota recesivo de ambos genes enmascara al otro gen o es epistático a sus efectos.
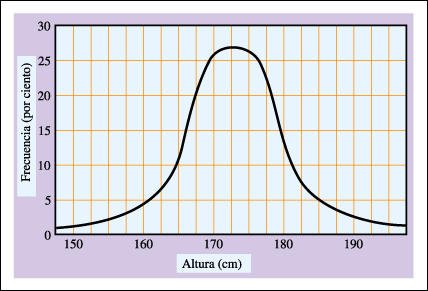
Distribución de altura de los hombres.
La altura es un ejemplo de herencia
poligénica; es decir está afectada por varios genes. La curva
de distribución de la altura tiene forma de campana, como se muestra
aquí, con la media o promedio cayendo habitualmente en el centro de la
curva.
Alteraciones cromosómicas
En los cromosomas pueden ocurrir
cambios que afecten su número o estructura. Estos cambios se clasifican
como alteraciones cromosómicas numéricas o alteraciones cromosómicas
estructurales, respectivamente.
Las alteraciones numéricas
pueden involucrar una dotación haploide completa o sólo algunos
cromosomas y, en general, se deben a fallas en la migración de los cromosomas
durante la meiosis o la mitosis. Por ejemplo, los organismos eucariotas diploides
presentan células o individuos haploides aunque, frecuentemente, esto
no constituye una situación anormal. Tal es el caso de los gametos y
de ciertas castas de abejas y hormigas que son haploides porque proceden de
huevos no fecundados. En genética vegetal, suelen obtenerse experimentalmente
organismos haploides y, de este modo, se consigue que ciertas variantes recesivas
se expresen siempre. Seleccionando artificialmente esas variantes, pueden construirse
ejemplares resistentes a diferentes factores del medio ambiente y luego, por
manipulación de la mitosis -empleando agentes que impiden la formación
del huso mitótico - obtener líneas puras de diploides homocigotas,
derivados de los haploides seleccionados.
Otras veces, la dotación cromosómica es superior a dos y, en este caso, los organismos son poliploides (triploides, 3n; tetraploides, 4n; pentaploides, 5n; etc.) lo que representa, a veces, una situación anormal. Sin embargo, los poliploides son muy frecuentes entre las plantas.
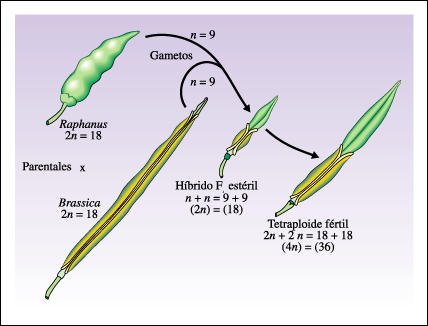
Origen del tetraploide fértil (4n = 36) entre la col, Brassica (2n = 18), y el rábano, Raphanus (2n = 18).
En otros casos, los cambios
en el número de cromosomas no afectan a una dotación completa
sino que involucran a uno o a unos pocos cromosomas. Por ejemplo, el síndrome
de Down está caracterizado por una trisomíaen el par 21 (tres
cromosomas del par 21) y el síndrome de Turner por una monosomíadel
cromosoma X (el complemento sexual integrado por un solo cromosoma X).
Las alteraciones estructurales se deben a rupturas cromosómicas que ocurren dentro de un cromosoma o entre cromosomas no homólogos. Una porción de un cromosoma puede perderse y sufrir una deleción, puede duplicarse, puede ser translocada a un cromosoma no homólogo, o puede invertirse. Los estudios hechos en los cromosomas gigantes de las larvas de Drosophila suministraron la confirmación visual de estos cambios, así como la evidencia final y concluyente de que los cromosomas son los portadores de las partículas de la herencia.
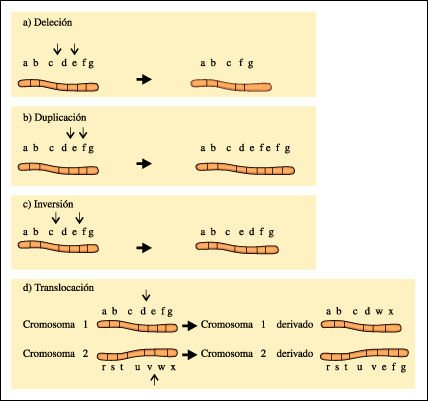
Alteraciones cromosómicas estructurales. a) Una porción de un cromosoma puede perderse, y sufrir una deleción; b) puede duplicarse; c) puede invertirse o d) puede ser translocada a un cromosoma no homólogo.


