Capítulo 18.
Desarrollo: la ejecución de un programa genético
El desarrollo es el proceso por
el cual un huevo fecundado se transforma en un organismo adulto. Este proceso
está controlado genéticamente, como lo indican su reproducibilidad
a través de las generaciones y la existencia de mutaciones que interrumpen
pasos específicos.
Un organismo adulto puede entenderse
como un arreglo ordenado de tipos celulares específicos. Los distintos
tipos celulares aparecen durante el desarrollo a través del proceso de
diferenciación celular, el cual implica la expresión diferencial
de genes específicos.
La expresión diferencial
de genes se logra a través de la regulación diferencial de esos
genes por proteínas llamadas reguladores maestros. Estas proteínas
pueden regular genes específicos de tipos celulares (músculo,
glóbulos rojos, piel) o pueden definir regiones anatómicas mayores
como los segmentos de los insectos.
Los reguladores maestros que
definen regiones específicas se denominan selectores; un grupo de células
contiguas que expresan un mismo selector constituyen un compartimiento.
A medida que las células
de un embrión se dividen, generalmente el número de compartimientos
aumenta, es decir, aparecen nuevas poblaciones de células que expresan
selectores específicos. Para que esto suceda, es necesario que se produzcan
divisiones asimétricas: divisiones en las que las células hijas
son diferentes de la célula progenitora.
En el desarrollo coherente de
un organismo, a medida que las poblaciones de células se vuelven más
heterogéneas, es decir, a medida que aparecen más y más
compartimientos, la ubicación espacial de estas regiones es clave. Existen
varios mecanismos que permiten la coordinación del desarrollo.
El proceso básico del
desarrollo embrionario temprano en la mosca D. melanogaster reside en el establecimiento
de la expresión de selectores -los genes maestros- que definen su plan
anatómico fundamental. Existen sólo cuatro rutas principales relativamente
independientes por las que se establece el plan anatómico en Drosophila.
En estas rutas, ciertos factores inician una cascada de eventos regulatorios,
activando genes cuyos productos regulan, a su vez, otra serie de genes, y así
se van estableciendo, en el embrión compartimientos cada vez más
pequeños en un patrón más detallado. La cascada culmina
con el establecimiento correcto de los patrones de expresión de los selectores
clave. El desarrollo de las diferentes estructuras es promovida por morfógenos
en función de su concentración. Mediante el establecimiento de
gradientes de concentración de morfógenos es posible la coordinación
armoniosa de las estructuras anatómicas.
La coordinación del desarrollo
también puede darse por medio de inducciones sucesivas. Una célula
o grupo de células emite una señal química que induce un
cambio en las células vecinas. Éstas, como respuesta a la señal,
producen una segunda señal que induce otro cambio en un tercer conjunto
de células. Esta cascada de inducciones también puede garantizar
el correcto ordenamiento de estructuras en el desarrollo. La embriogénesis
de vertebrados ilustra muy bien ese mecanismo, de la misma manera que lo hace
la formación de la vulva del gusano C. elegans.
El estudio de los genes del
desarrollo en diversos organismos ha conducido al descubrimiento de semejanzas
entre grupos diversos hasta el momento soslayadas. Muchos de los genes del desarrollo
están tan conservados que pueden ser transplantados de un organismo a
otro sin perjuicio para el desarrollo del organismo receptor. Esto conduce a
preguntarse qué cambios fueron los que operaron durante la evolución
en la Tierra, modificaron los programas de desarrollo y generaron la diversidad
anatómica observable.
El desarrollo de las plantas
es significativamente distinto del de los animales, debido fundamentalmente
a su carácter sésil. Se han encontrado numerosos genes que controlan
el crecimiento vegetal.
Uno de los aspectos mejor conocidos
del desarrollo vegetal es la serie de mecanismos que determinan la identidad
de los órganos florales.
La
diferenciación: la expresión diferencial de genes
En el organismo adulto existen
arreglos ordenado de tipos de células específicas. Cada tipo celular
está caracterizado por la acción diferencial de no una, sino muchas
proteínas específicas; para que esas proteínas se sinteticen,
ciertos conjuntos de genes deberían encenderse concurrentemente mientras
que otros estarían reprimidos.
Muchos de los experimentos iniciales
sobre expresión diferencial se realizaron en líneas celulares
-poblaciones homogéneas de células que se mantienen vivan fuera
del animal en medios de cultivo-. Existen líneas celulares de distintas
identidades, como las musculares. Inicialmente se observó que ciertos
genes, entre ellos myoD y myf5, estaban activos exclusivamente en líneas
celulares musculares. Cuando se forzó experimentalmente la expresión
de estos genes en líneas celulares no musculares, esas células
se diferenciaron en músculo. Se comprobó que esos genes codificaban
factores de transcripción que actuarían sobre otros genes, es
decir, que inducirían la diferenciación celular mediante la regulación
de una segunda línea de genes específicos. Por esta razón,
a esos factores de transcripción se les dio el nombre de reguladores
maestros y, a los genes que los codifican, el de genes maestros. Tal como una
llave maestra de una cañería cierra el paso a muchas otras válvulas
subsidiarias, los reguladores maestros controlan la expresión de numerosos
genes subalternos.
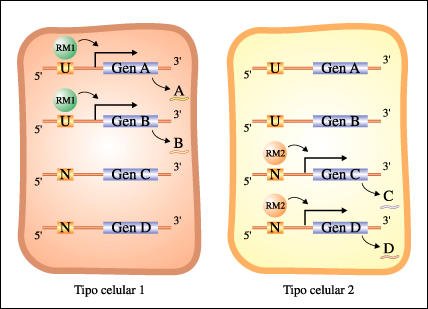
En el ejemplo de la figura siguiente, el tipo celular 1 está caracterizado por la presencia de los productos de los genes A y B. El tipo celular 2 está caracterizado por la presencia de los productos de los genes C y D. Ambos tipos celulares contienen copias de todos los genes. El Regulador Maestro 1 (RM1) se une a los sitios "U" y activa los genes que contienen ese sitio en su región promotora. El Regulador Maestro 2 (RM2) se une a los sitios "N". Las células de tipo 1 contienen el regulador RM1 (una proteína). Por lo tanto, todos los genes con sitios promotores "U" son activados. Como carecen de la proteína reguladora RM2, los genes con regiones promotoras con sitios "N" no son activados. Para el tipo celular 2 se produce la situación inversa.
La diferenciación celular es el resultado de la expresión diferencial de genes bajo el control de reguladores maestros.
En ausencia de reguladores maestros funcionales, se reprime la expresión
de numerosos genes subalternos mientras que si el regulador maestro está
presente, estos genes subalternos son forzados a expresarse.
En conclusión, la clave
por la que se generan distintos tipos celulares reside, entonces, en que distintas
células contienen diferentes proteínas reguladoras específicas.
Selectores y compartimientos
Los genes homeóticos
son genes maestros que no determinan tipos celulares, sino regiones anatómicas
completas, por ejemplo, segmentos. Cada gen homeótico dirige el desarrollo
de un segmento o grupo de segmentos en particular mediante la regulación
de infinidad de genes subalternos.
Se demostró que los genes
homeóticos no se expresan en una única célula sino en bloques
de células que coinciden con segmentos o grupos de segmentos y que, efectivamente,
regulan la expresión de otros genes. Sus productos -los reguladores maestros-
son factores de transcripción.
Cuando uno de estos genes homeóticos
es activado fuera de su dominio normal de acción, activa sus genes subalternos
en el lugar que ha invadido, forzando la aparición de características
anatómicas propias de su dominio de origen.
Los genes maestros que gobiernan
regiones anatómicas se llaman selectores. Los genes homeóticos
reciben pues el nombre de selectores homeóticos.
Por otra parte, un grupo de
células que expresan un selector particular se define como compartimiento.
Por ejemplo, las células que forman una pata en las moscas, expresan
un selector homeótico característico del primer segmento torácico
(en los insectos, las patas se insertan en el tórax).
En la siguiente figura se pueden observar los efectos de las mutaciones en dos grupos de genes homeóticos de Drosophila.
A la izquierda, mosquita normal con un par de alas y un par de balancines y a la derecha, mosquita mutante con dos pares de alas y sin balancines
Dos estrategias que generan
células diferentes
Es posible que se produzcan
divisiones celulares en las que las células hijas sean diferentes de
la célula progenitora. Células inicialmente equivalentes pueden
ser conducidas a expresar selectores distintos por comunicación entre
las células. Como todas las células del organismo derivan, en
última instancia, del huevo fertilizado, en algún momento deben
producirse divisiones que produzcan dos células diferentes: una con un
regulador maestro y otra sin éste; son divisiones asimétricas.
Existen varios mecanismos que
generan células o poblaciones de células diferentes a partir de
una célula. En uno de estos mecanismos, una célula puede tener
una distribución asimétrica de moléculas en su citoplasma.
Por ejemplo, un extremo puede contener el RNA mensajero correspondiente a un
regulador maestro, y el otro extremo carecer de él. Si una división
celular separa estas dos fracciones de citoplasma en dos células hijas,
se produce una división asimétrica y se obtienen, naturalmente,
dos células diferentes. Este mecanismo simple recibe el nombre de segregación
de factores citoplasmáticos.
Las divisiones celulares asimétricas
son aquellas en las cuales las dos células hijas son diferentes -en su
estructura o en sus propiedades biológicas- de la célula progenitora.
Existen varios mecanismos que dan como resultado divisiones celulares asimétricas:
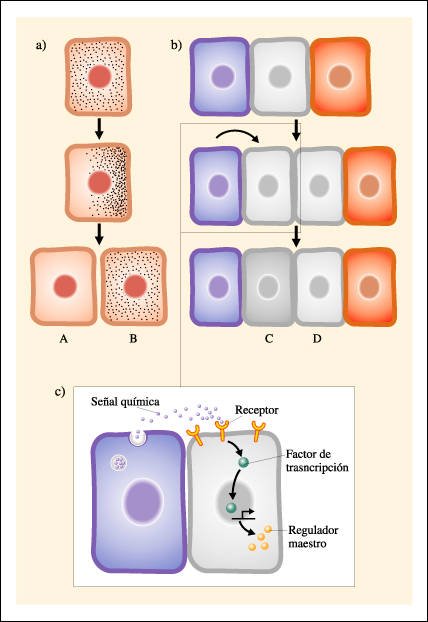
Mecanismos por los cuales se producen divisiones celulares asimétricas
a) Segregación de factores
citoplasmáticos: La célula madre contiene ciertas sustancias distribuidas
asimétricamente en su citoplasma. Al producirse la citocinesis, cada
célula hija (A y B) lleva sustancias diferentes. Éstas pueden
ser factores de transcripción o moléculas de mRNA que codifican
factores de transcripción. En consecuencia, cada célula hija expresará
un grupo diferente de genes. b) Comunicación celular: Las células
hijas pueden ser idénticas en un principio y distinguirse luego (en células
C y D, por ejemplo) por la acción de sustancias químicas que actúan
como señales exógenas.
En la figura, en un primer momento,
la célula de color gris está rodeada por dos células, azul
y roja. La célula gris se divide y da dos células hijas iguales
(grises). La célula hija de la izquierda recibe de su vecina (azul una
señal química. En principio, las dos células hijas grises
son capaces de responder a la señal, pero sólo lo hará
aquella que se encuentre físicamente cerca de la fuente de la señal
química y pueda así entrar en contacto con esa señal. Las
señales exógenas pueden ser producidas por otras células
o por el medio circundante, como la matriz celular de tejidos vecinos. c)Detalle
del mecanismo celular que traduce una señal externa en una respuesta
interna. La señal es una molécula con una estructura tridimensional
específica. La membrana de la célula ubicada a la derecha posee
un receptor, una proteína con una porción extracelular y otra
intracelular. La porción extracelular posee afinidad específica
por la señal (similar a la afinidad de una enzima por su sustrato). Cuando
el receptor se une a la señal, se operan cambios en la porción
intracelular, la cual es entonces capaz de activar la expresión de un
gen maestro que redundará en la síntesis de un regulador maestro.
Por otra parte, una división
o serie de divisiones celulares puede producir un conjunto de células
idénticas, que luego sean estimuladas diferencialmente por su entorno.
El entorno de las células hijas puede ser diferente, por ejemplo, por
tener distintas células vecinas. Las células en principio idénticas
reciben distintas señales del exterior que provocan la activación
o inactivación de un regulador maestro específico. A este mecanismo,
muchas veces llamado inducción, también se lo denomina comunicación
celular.
La coordinación de los
procesos de desarrollo
En el desarrollo coherente de
un organismo, es fundamental la coordinación de los procesos de desarrollo
y la ubicación espacial de compartimientos.Existen
varios mecanismos que permiten la coordinación del desarrollo.
Es posible analizar cómo
se produce esta coordinación en algunos organismos en los que se han
producido grandes avances en el estudio de su genética; esto ha permitido
identificar los procesos clave.
Un ejemplo es el de Caenorhabditis
elegans, un gusano cilíndrico que pertenece al grupo de los nematodos.
Las ventajas experimentales de este organismo incluyen su pequeño tamaño,
su ciclo de vida corto, su fácil manejo, y el hecho de que su cuerpo
es transparente. Esta última característica permite el examen
directo del proceso de desarrollo.
Los nematodos tienen otra característica
curiosa: el número de núcleos somáticos del adulto es invariable
y característico de la especie: por ejemplo, C. elegans tiene 959 núcleos
somáticos. Además, las posiciones relativas de las células
durante el desarrollo también es invariable de individuo a individuo,
de modo que todo el proceso de desarrollo, desde el cigoto al adulto, puede
esquematizarse como un árbol genealógico celular.
El árbol genealógico
de las células de C. elegans se conoce en forma completa.
En el árbol de la figura, cada bifurcación representa una división celular. No todas las divisiones se muestran. Los globos de color rosa denotan conjuntos celulares.
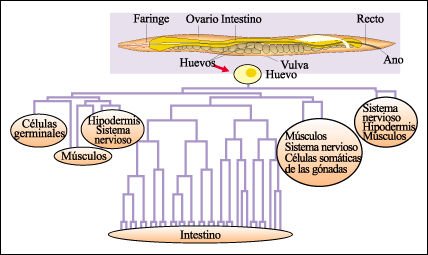
El estudio del desarrollo de C. elegans permitió seguir el camino de cada una de sus células a través de las sucesivas divisiones celulares, desde la primera hasta la número 962 y así trazar su "árbol genealógico celular".
En C. elegans, esta cadena de divisiones mitóticas tan finamente coreografiada
se inicia con el huevo fecundado, que se encuentra dentro de una dura cáscara
externa. Las tres primeras divisiones producen un embrión de cuatro células,
cada una con un destino embriológico determinado. En la primera división
se produce la segregación de factores citoplasmáticos que mencionamos
recientemente y se obtiene, por lo tanto, dos células con contenidos
distintos. Una de ellas, la célula P, dará lugar al polo posterior
del embrión, mientras que la otra, la célula AB, dará lugar
al polo anterior.
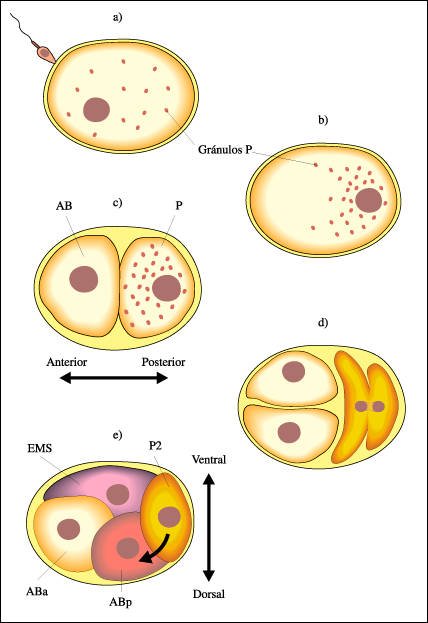
En el desarrollo temprano de C. elegans, la complejidad celular aumenta a través de varios mecanismos
El gameto femenino de C. elegans
contiene una cubierta muy dura que limitará los movimientos de las células
que se formarán en su interior luego de la fecundación.
a) El espermatozoide penetra por un agujero a través de la cáscara
y se produce la fecundación .
b) En el punto de entrada del espermatozoide, se organiza un eje de división
celular y se concentran unos gránulos oscuros llamados "gránulos
P". Los gránulos P promueven el desarrollo de estructuras posteriores.
c) La primera división celular conlleva la segregación de factores
citoplasmáticos y produce dos células distintas: la célula
P (que contiene gránulos P) y que genera las estructuras posteriores,
y la célula AB, que produce las estructuras anteriores. Esta primera
división, por lo tanto, define el eje antero-posterior del animal.
d) La célula P se divide en dos células distintas llamadas EMS
y P2. La célula AB se divide en dos células equivalentes pero,
debido al espacio limitado dentro de la cáscara del huevo, sólo
una de ellas estará físicamente en contacto con la célula
P2. Esta última produce una señal (representada por una flecha)
que permanece adherida a su membrana citoplasmática y promueve cambios
en su célula vecina.
Así, una célula
descendiente de AB queda en contacto con la célula P2 y pasa a convertirse
en la célula ABp. La otra célula que deriva de AB es la célula
ABa que no tiene contacto con P2. ABa y ABp dan lugar a estructuras diferentes;
sus destinos dependen de la señal producida por P2. ABp define la región
dorsal del embrión, por lo que este proceso de comunicación celular
también define, en parte, el eje dorso-ventral del animal.
La célula P también
se divide asimétricamente por segregación de factores citoplasmáticos.
La célula AB, sin embargo, da lugar a células con potencialidades
idénticas pero que pronto adquieren identidades distintas al recibir
señales de su entorno. Las limitaciones de espacio dentro de la dura
cáscara del huevo obligan a estas células a ocupar determinadas
posiciones con respecto a las demás. Las células se encuentran,
por lo tanto, expuestas a diferentes señales químicas y, en consecuencia,
asumen identidades y destinos embriológicos diferentes. Este es un claro
ejemplo de división asimétrica por comunicación celular.
Es notable que, las diferentes
células del embrión de C. elegans tienen destinos embriológicos
específicos desde temprano. Si cualquiera de estas células es
destruida, ninguna de las otras podrá generar las estructuras que éstas
hubieran generado. Esto contrasta con lo que sucede en los embriones de muchos
otros organismos, los cuales producen poblaciones de células equivalentes
que adquieren un destino embriológico más tarde en su desarrollo;
en esos casos, la destrucción de una célula es compensada por
sus vecinas. El desarrollo temprano del gusano C. elegans está dado,
entonces, por dos factores. Por un lado, las limitaciones de espacio impuestas
por la cáscara del huevo obligan a las células a ocupar posiciones
determinadas unas con respecto a otras por lo que cada una recibe distintas
señales químicas. Por otro lado, las divisiones celulares asimétricas
asignan un destino determinado a cada célula en cada posición.
El desarrollo de la vulva del
gusano C. elegans comienza cuando el embrión ya está compuesto
por numerosas células e ilustra otro mecanismo por el cual se genera
una coherencia espacial en la disposición de las células. La vulva
de C. elegans es un órgano sencillo: un pequeño orificio por donde
entra el esperma y salen los huevos. Está compuesta por dos tipos celulares
específicos: el tipo 1 que conecta la gónada con el exterior,
y el tipo 2, que brinda protección al primer tipo. En total son 22 células,
derivadas en principio de un conjunto de tan sólo tres células
ubicadas en el lado ventral del gusano. Para que el órgano se desarrolle
con su estructura correcta, las células precursoras deben tener una disposición
que denominaremos 2-1-2 (una célula precursora del tipo 1 rodeada, a
cada lado, por una célula precursora del tipo 2). Esta sencilla disposición
se establece por un sistema de comunicación de señales entre células.
Los dos tipos celulares mencionados
derivan de un conjunto de células epidérmicas equivalentes, todas
con el potencial de diferenciarse en tipos 1 ó 2. El desarrollo de la
vulva es gatillado por una célula que no formará parte de ese
órgano conocida como célula ancla.
Si la célula ancla es
destruida, por ejemplo, por ablación con láser, la vulva no se
forma en absoluto.
La célula ancla produce una proteína señal que es liberada
al medio extracelular por exocitosis. La célula epidérmica más
cercana a la célula ancla adquirirá, por acción de la proteína
señal, la identidad de tipo 1. Esta célula, a su vez, producirá
una segunda señal, que difundirá desde la célula tipo 1,
induciendo a sus dos vecinas a convertirse en células del tipo 2. El
resultado de esta concatenación en el tiempo de señales de acción
a corta distancia es el patrón 2-1-2 de la vulva.
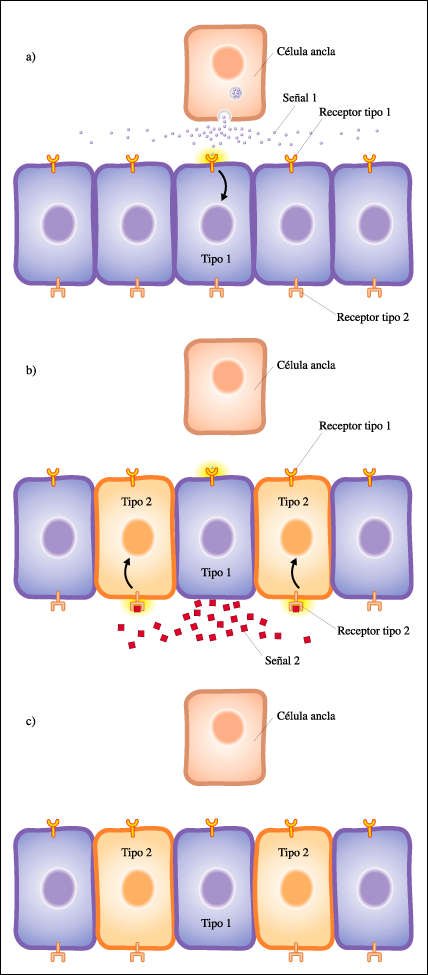
La vulva de C. elegans presenta un patrón de tipos celulares que surge como consecuencia de comunicaciones celulares sucesivas.
a. En el epitelio ventral existe
un conjunto de células con la capacidad de diferenciarse en todos los
tipos celulares presentes en la vulva. Esta capacidad se expresa en la presencia
de receptores en sus membranas. Todas las células contienen un tipo de
receptor proteico para una señal (señal 1, representada por círculos)
producida por la célula ancla. Esta señal difunde libremente en
el espacio intercelular y su concentración va disminuyendo con la distancia
a la fuente donde se produce esa señal. La célula epitelial más
cercana a la célula ancla estará expuesta a concentraciones suficientemente
altas de la señal como para que ésta pueda interactuar con su
receptor de membrana específico y activarlo. Cuando el receptor es activado,
promueve la actividad de un regulador maestro específico de células
tipo 1. Entonces, los genes característicos de este tipo celular son
activados, y los del tipo 2 son reprimidos o no son activados.
b. Entre los genes activados,
se encuentra uno que codifica una segunda señal (señal 2, representada
por cuadradros), la cual es transportada hacia la membrana y liberada por exocitosis.
Entre los genes reprimidos se encuentra el que codifica un receptor para esa
señal, por lo que la célula de tipo 1 produce una señal
química pero ella misma es incapaz de responder a esa señal. La
señal producida por la célula tipo 1 migra por difusión,
disminuyendo su concentración, de modo que sólo las dos células
adyacentes encuentran una concentración de señal suficientemente
alta como para que se pueda activar el receptor específico. En estas
células, la activación del receptor de tipo 2 conduce a la acción
del regulador maestro específico del tipo celular 2. Este regulador maestro
no es activado en la célula 1 porque ésta carece del receptor
capaz de interactuar con la señal.
c. Mediante la producción
sucesiva de señales entre células, se consigue el patrón
2-1-2 que permitirá el desarrollo armonioso de la vulva. Este es un ejemplo
sencillo de cómo la diferenciación celular es coordinada en el
espacio y produce así un conjunto coherente.
La respuesta a una señal
no es necesariamente de inducción de un tipo celular: también
hay señales inhibitorias, y en abundancia. Es posible, a veces, que entre
los precursores de la vulva de C. elegans, dos células lleguen a adquirir
la identidad de tipo 1. Cuando esto sucede, comienza una competencia entre las
dos por la supremacía mediante la producción de señales
inhibitorias. Este mecanismo también garantiza la organización
2-1-2, impidiendo situaciones del tipo 2-1-1-2 y, además, ilustra el
hecho de que no todas las señales son estimulatorias, sino que la inhibición
también juega un papel importante en el desarrollo.
Cómo "se construye"
una mosca: cascadas genéticas en Drosophila
En los últimos veinte
años, la mosca de la fruta, D. melanogaster se ha convertido en el organismo
modelo para el estudio genético del desarrollo.
El proceso básico del
desarrollo embrionario temprano reside en el establecimiento de la expresión
de los selectores -los genes maestros- que definen el plan anatómico
fundamental de la mosca. Estos selectores son muchos pero el estudio del desarrollo
temprano permitió comprender que existen sólo cuatro rutas principales
relativamente independientes por las que se establece el plan anatómico
de la mosca. En cada una de estas rutas, un pequeño grupo de factores
inician una cascada de eventos regulatorios, activando una serie de genes maestros;
los productos de estos genes regulan, a su vez, otra serie de genes maestros,
cuyos productos actúan sobre otra serie de genes subalternos. En cada
paso, se establecen, en el embrión, compartimientos cada vez más
pequeños en un patrón más detallado. La cascada culmina
con el establecimiento correcto de los patrones de expresión de los selectores
clave.
Existen tres rutas de desarrollo
a lo largo del eje antero-posterior: dos de ellas interactúan de modo
que dan forma a la región central, segmentada, del embrión, mientras
que la tercera define los extremos no segmentados, tanto anterior como posterior.
La cuarta ruta de desarrollo define las diferencias a lo largo del eje dorso-ventral.
Las dos primeras cascadas de
activación de genes mencionadas coordinan la formación de la región
central del embrión de Drosophila. Esta región se caracteriza
por estar dividida en segmentos. Los segmentos tienen un diseño básico
común, pero que presenta variaciones de un segmento a otro. Un conjunto
de genes es responsable de generar el patrón reiterativo, es decir segmental,
del cuerpo de Drosophila. Este conjunto de genes recibe el nombre de genes de
polaridad de segmento. Son los responsables de mantener los límites entre
los segmentos y son también necesarios para establecer los patrones estructurales
comunes a todos los segmentos. Cuando un gen de polaridad de segmento muta,
sobreviene un defecto que se repite en cada segmento. Algunos de los genes de
polaridad de segmento, pero no todos, son factores de transcripción.
Otro conjunto de genes es responsable de las variaciones que diferencian un
segmento del otro. Estos son los selectores homeóticos. Cuando un selector
homeótico muta, el resultado fenotípico es que un segmento asume
el aspecto de otro.
Antes de ser fertilizado, un
gameto femenino de Drosophila, como los de posiblemente todas las especies,
contiene abundante material citoplasmático provisto por la madre. Entre
estas moléculas se encuentran también una serie de moléculas
de mRNA que encierran las primeras instrucciones del desarrollo del embrión.
Esta circunstancia implica que los primeros pasos del desarrollo de una mosca
no están dirigidos por sus propios genes, sino por los genes de su madre.
Genes con este tipo de efecto reciben el nombre de genes de efecto materno.
Los productos proteicos de ciertos
genes maternos, denominados morfógenos, se encuentran asimétricamente
distribuidos en el citoplasma del huevo. Hacia el extremo que luego será
el anterior, existen altas concentraciones de una proteína llamada Bicoid.
Hacia el extremo posterior, hay altas concentraciones de una proteína
llamada Nanos. La concentración de ambas proteínas decrece al
alejarse de los extremos, formando gradientes de concentración.
La acción de los morfógenos
y su traducción en un patrón de expresión génica
nos da un ejemplo concreto de cómo la activación localizada de
reguladores maestros puede coordinarse en el espacio y, así, dar un todo
coherente. Los morfógenos Bicoid y Nanos esencialmente crean un sistema
de coordenadas con valores precisos de concentraciones en el eje antero-posterior.
Estos valores son leídos por sistemas que expanden y elaboran las diferencias
iniciales, pero la polaridad está dada desde el principio por el gradiente
de concentraciones, y es ésta la que establece la coordinación
de la anatomía del organismo.
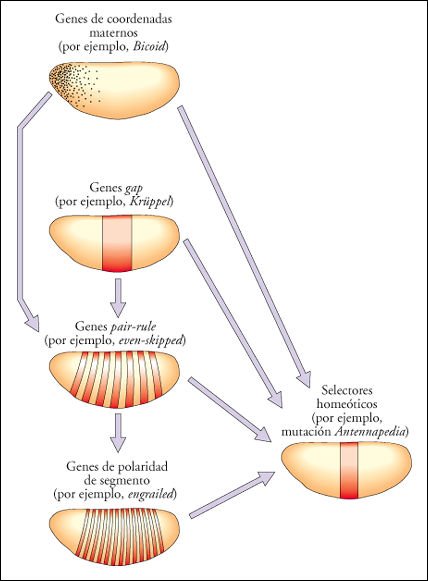
Otra observación importante es la organización jerárquica de los procesos del desarrollo. En la cúspide de la jerarquía que analizamos anteriormente se encuentran los dos morfógenos Bicoid y Nanos, le siguen los genes gap, los pair-rule, los selectores homeóticos y los genes de polaridad de segmento. Esta organización jerárquica sigue extendiéndose hacia abajo. No podemos saber hasta dónde se extiende la cascada regulatoria, pero sí sabemos que en el nivel más bajo se encuentra el conjunto de genes estructurales como los que codifican para proteínas como la actina, la queratina y la hemoglobina. Estos genes ejecutan la diferenciación celular, dándole a cada célula y tejido las propiedades físicas, químicas y biológicas que los caracterizan.
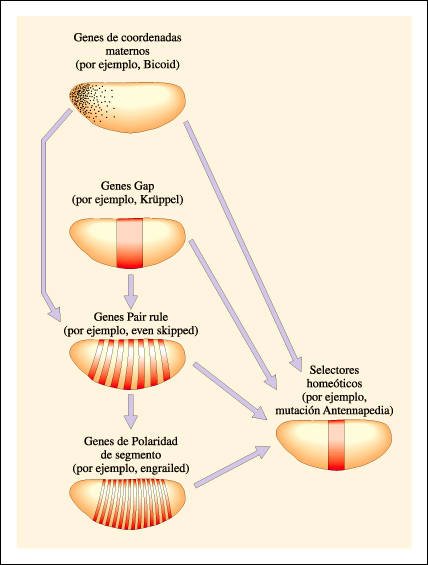
Cascada de regulación génica en la embriogénesis del eje antero-posterior de Drosophila .
El plan anatómico de
los insectos se caracteriza, en el eje antero-posterior, por estar dividido
en segmentos (dependientes de la actividad de los genes de polaridad de segmento).
Cada segmento muestra especializaciones propias que dependen de la acción
de los selectores homeóticos.
El establecimiento del plan
anatómico es el resultado de una cascada de reguladores génicos,
organizados jerárquicamente. Al tope de la jerarquía se encuentran
los morfógenos depositados por los tejidos de la madre. Los genes que
codifican estos morfógenos, y muchos otros genes maternos necesarios
para la acción de los primeros, reciben el nombre colectivo de genes
de coordenadas maternos. Estos genes son necesarios para la correcta expresión
de los genes gap, los cuales se expresan en bloques o dominios contiguos a lo
largo del embrión.
A su vez, combinaciones de proteínas
codificadas por genes gap y por los genes de coordenadas maternos, promueven
la expresión de genes pair-rule en siete bandas discretas. Los genes
pair-rule son numerosos y sus productos de expresión se solapan. Combinaciones
particulares de proteínas pair-rule promueven la expresión de
genes de polaridad de segmento.
Los selectores homeóticos
son activados en sus dominios específicos por la acción combinada
de los productos de todos los demás genes. Muchos de estos genes, además,
regulan a otros dentro de la misma categoría, incluso a sí mismos
(autorregulación).
Desarrollo temprano en vertebrados:
inducciones sucesivas
La mayor parte de los experimentos
llevados a cabo en vertebrados han sido realizados en anfibios porque sus huevos
son grandes, no tienen cáscara y son de fácil manipulación.
En aves, reptiles y mamíferos, en cambio, el embrión se encuentra
inaccesible dentro de la cáscara o del útero.
El desarrollo de un anfibio
comienza antes de la fecundación. El gameto femenino tiene una distribución
asimétrica de sustancias, la cual es incluso visualmente evidente: una
mitad es incolora y la otra contiene un pigmento oscuro.
Poco después de la fertilización
aparece, en la zona donde las mitades se juntan, pero sólo en un costado,
una tercera zona: una delgada franja llamada medialuna gris. Las tres zonas
contienen diferentes sustancias que incluyen mRNA específicos. Al producirse
las primeras divisiones celulares, se producen células diferentes debido
al mecanismo de segregación de factores citoplasmáticos.
Cada una de las zonas establecidas
en el gameto femenino dará lugar a diferentes estructuras del embrión.
Las células que reciben el material incluido en la medialuna gris son
especiales: ellas no contribuirán físicamente a muchas estructuras,
pero su presencia es indispensable para el desarrollo de muchas de las partes
del embrión.
El material de la medialuna gris es capaz de inducir el desarrollo de muchas de las estructuras anatómicas, a pesar de que las células que lo contienen no forman parte en sí de esas estructuras.
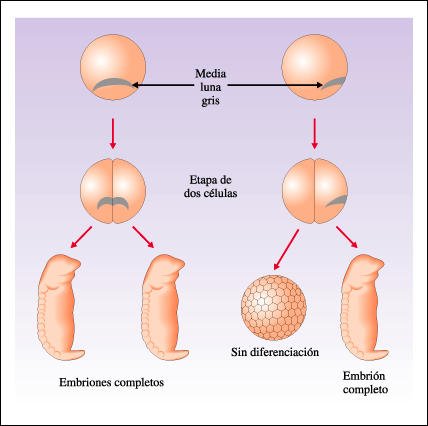
Experimento que demuestra la importancia del citoplasma de la media luna gris en el desarrollo.
La importancia
del citoplasma de la media luna gris en el desarrollo fue demostrada al separar
las dos células formadas por la primera segmentación del huevo.
Cuando el huevo de la izquierda se dividió, la mitad del citoplasma de
la media luna gris pasó a cada una de las dos células nuevas.
Cuando estas células fueron separadas una de otra, cada una formó
un embrión completo. La primera segmentación del huevo de la derecha
dio como resultado que todo el citoplasma de la media luna gris quedara en una
célula. Cuando estas células fueron separadas, la que no tenía
media luna no se desarrolló.
Las células que contienen el material de la medialuna gris -que forman el centro de Nieuwkoop- producen ciertas señales químicas que promueven cambios en sus vecinas. Como resultado de esos cambios, un conjunto de células -que forman el organizador de Spemann - comienza a expresar ciertos genes entre los que se encuentran aquellos que codifican una segunda generación de señales y "organizan" así a las células vecinas en un embrión casi completo.
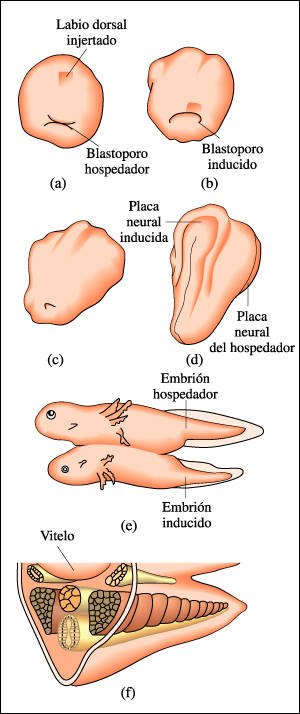
Experimento que muestra la actividad del organizador.
La actividad del organizador se puede comprobar en el sigyuiente experimento:
a) El labio dorsal del blastoporo de un embrión de anfibio se injerta
en otro embrión. b) El labio dorsal injertado induce la formación
de un segundo blastoporo, a través del cual el tejido injertado se mueve
al interior del embrión durante la gastrulación. c) A medida que
prosigue el desarrollo, se forman dos conjuntos de estructuras embrionarias,
y d) se produce un embrión doble.
La señal emitida por
las células del organizador de Spemann tiene efectos variados dependiendo
de qué células la reciban. La epidermis que recibe la señal
del organizador sufre cambios espectaculares, generando una estructura en forma
de tubo, la cual posteriormente formará la médula espinal del
sistema nervioso. Como este tubo -el tubo neural- es obviamente más largo
que ancho, define un eje: uno de sus extremos define la cabeza y el otro la
cola del embrión. Su posición también define el lado dorsal
del embrión. Así, la posición del organizador y de las
moléculas que produce establecen la polaridad en dos ejes en el embrión.
Parte de las células
del organizador forman más tarde la notocorda, una masa apretada de células
de forma cilíndrica que corre a lo largo del eje antero-posterior, justo
por debajo del tubo neural. La notocorda también produce señales.
Estas señales inducen cambios en la parte ventral del tubo neural, la
cual, a su vez, produce otras señales que inducen cambios en algunas
células de la parte dorsal del tubo neural. Mediante estos procesos,
el tubo neural es dividido en varias zonas con propiedades y destinos distintos.
La cascada continúa y, en cada paso, un conjunto de células es
dividido en varios conjuntos con identidades distintas, posicionados en el lugar
adecuado con respecto a las demás estructuras. Cada zona expresa reguladores
maestros distintos y constituye, entonces, un compartimiento. El mecanismo que
garantiza la coordinación espacial de las estructuras está basado
fundamentalmente en la emisión de señales químicas que
actúan a cortas distancias.
El proceso de desarrollo de
los vertebrados puede ser descripto de manera simplificada como una cascada
de inducciones jerárquicas en la que, a cada paso, se aumenta el número
de reguladores maestros encendidos. La diferencia inicial es una distribución
asimétrica de factores citoplasmáticos.
Conservación y evolución
en los genes del desarrollo
La morfología final de
un organismo es el resultado de la ejecución de su programa genético
de desarrollo. Los cambios en la morfología que vemos en distintas especies
se deben a cambios en ese programa de desarrollo. Entender la genética
del desarrollo significa dar luz a una de las áreas más intrigantes
de la biología evolutiva que plantea numerosos interrogantes: ¿Cómo
surgieron las diferentes estructuras anatómicas que vemos hoy? ¿Qué
cambios en el desarrollo pueden hacer que un miembro crezca como una pata y
no como una aleta? ¿Qué genes sufrieron mutaciones o qué
relaciones entre genes se modificaron en el curso de la evolución y dieron
lugar a un ala, una mandíbula o un ojo? La investigación de la
evolución a nivel morfológico ha tenido un enorme impulso por
parte de la genética del desarrollo en los últimos años;
una de sus metas más ambiciosas es entender el surgimiento de las llamadas
novedades anatómicas.
La organización jerárquica
de los programas genéticos de desarrollo es sumamente atractiva en el
sentido que sugiere que mutaciones puntuales pueden conducir a reordenamientos
profundos del plan anatómico de un organismo. Es suficiente con que mute
un gen regulador maestro en lo alto de la jerarquía para que una multitud
de genes subalternos modifique su acción. Las mutaciones homeóticas
fueron siempre favoritas entre los biólogos por su posible efecto pleiotrópico
sobre muchos otros genes. Para establecer cómo han cambiado los programas
genéticos en el curso de la evolución, es necesario realizar estudios
comparativos de esos programas en distintos organismos. Esta tarea no es sencilla,
por el simple hecho de que, para entender el programa genético es necesario
estudiar cientos de genes, y el estudio de cada uno de ellos requiere un enorme
esfuerzo. Esta es la razón por la cual sólo unos pocos organismos
son conocidos con cierto detalle a nivel genético. Afortunadamente, la
biología molecular nos da una mano allí donde la genética
encuentra su límite.
Uno de los primeros estudios
del desarrollo a nivel molecular, ahora paradigmático, fue el realizado
a principios de la década de 1980. Se sabía entonces que los selectores
homeóticos se encontraban agrupados en paquetes en un cromosoma de Drosophila.
Mediante hibridaciones con sondas
moleculares, los investigadores encontraron una corta secuencia de DNA que se
repetía, casi sin variación en todos los selectores homeóticos;
la llamaron homeobox. Se descubrió más tarde que el homeobox expresa
una secuencia de aminoácidos capaz de unirse al DNA y así regular
la transcripción.
No sólo todos los animales
contienen gran variedad de genes con secuencias homeobox, sino que todos ellos
tienen selectores homeóticos. Los estudios genéticos en ratón,
el gusano C. elegans, e insectos demuestra que la función de los selectores
homeóticos de establecer regiones en el eje antero-posterior está
fabulosamente conservada a lo largo de la evolución.
Si muchos de los genes que componen
los programas genéticos son los mismos en distintas especies, ¿qué
hace diferentes a estos programas de manera que produzcan formas tan disímiles?
En primer lugar, el hecho de que muchos genes estén conservados no significa
que todos los estén. En segundo lugar, hay evidencias que parecen indicar
que las diferencias entre los programas genéticos de desarrollo no yacen
fundamentalmente en los elementos de esos programas (los genes) sino en cómo
éstos interactúan unos con otros. Un factor clave parece ser cuándo
y dónde se expresan los genes del desarrollo. Por ejemplo, todos los
artrópodos tienen los mismos ocho selectores homeóticos, pero
los crustáceos y los insectos difieren en cuanto a cuáles y cuántos
segmentos expresan determinados selectores homeóticos. Es posible que
buena parte de la evolución morfológica, cuyo resultado es la
variedad de formas que vemos a nuestro alrededor, sea el resultado de mutaciones
en las regiones promotoras (regulatorias) de los genes y no en los genes mismos.
Desarrollo de plantas
Así como ocurrió
en el caso de los animales, los investigadores de la genética del reino
vegetal se han dedicado al estudio de algunos organismos modelo en los cuales
se concentra la mayor parte de las preguntas. Uno de los modelos más
estudiados es una pequeña planta llamada Arabidopsis thaliana, escogida
por su pequeño genoma y su fácil cultivo. Sin embargo, por razones
comerciales, se utilizan también otros modelos tradicionales como el
maíz y el tabaco.
El cigoto de Arabidopsis se
divide varias veces hasta formar un embrión con dos polos: uno dará
origen al meristema del tallo y otro al de la raíz. El embrión
tiene, además, otras estructuras como los cotiledones, que son las primeras
hojas con las que la plántula obtendrá su energía del Sol.
Ya la primera división celular de Arabidopsis es asimétrica, tanto
en lo que se refiere a la morfología de sus células como a los
genes que expresan, pero no se conocen cuáles son los mecanismos que
establecen estas asimetrías. Se han identificado a varios genes que participan
en el desarrollo embrionario.
Los meristemas son los grandes
arquitectos de la estructura de las plantas. Cuando las células de un
meristema de tallo se dividen, algunas de las células hijas se diferencian
en estructuras particulares, mientras que otras permanecen indiferenciadas.
Los meristemas de tallos y raíces no son los únicos meristemas existentes. En Arabidopsis, en cierto momento de su desarrollo, se produce un meristema de inflorescencia, el cual genera hojas de morfología particular, y una serie de meristemas de inflorescencia secundarios. Los meristemas de inflorescencia, tarde o temprano, adquieren una nueva identidad y dan lugar a flores en lugar de hojas y internudos, como ocurre en el crecimiento vegetativo. Cuando esto sucede se los llama, naturalmente, meristemas florales. La transición de un tipo de meristema a otro está mediada por distintos genes que controlan la identidad del meristema.
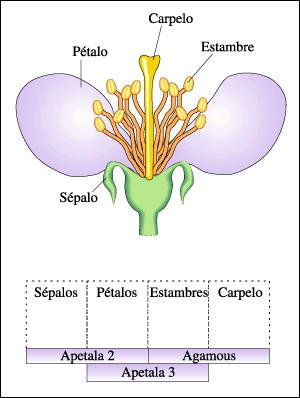
Genes homeóticos de las plantas que establecen la identidad de los órganos florales.
Existen cuatro órganos florales en forma de anillos concéntricos,
de afuera hacia adentro: sépalos, pétalos, estambres y carpelo.
La identidad de cada pieza está establecida por combinaciones de reguladores
maestros. Las células que sólo expresan el gen Apetala2 se desarrollan
en forma de sépalos. Las células que acumulan los productos de
los genes Apetala2 y Apetala3 siguen la ruta del desarrollo que culmina en pétalo.
Aquellas que combinan Apetala3 con Agamous se desarrollan como estambres y,
las que sólo contienen el regulador maestro codificado por el gen Agamous
forman el carpelo. Si alguno de estos patrones de expresión cambia, también
cambiarán las células que expresan ciertas combinaciones y sobrevendrán
transformaciones homeóticas.


