Capítulo 21.
Las bases genéticas de la evolución
La genética de poblaciones es una síntesis
de la teoría darwiniana de la evolución con los principios de
la genética mendeliana. Para el genetista de poblaciones, una población
es un grupo de organismos que se cruzan, definidos y unidos por su reservorio
génico. La evolución es el resultado de los cambios acumulados
en la composición del reservorio génico.
La amplitud de la variabilidad
genética en una población es un determinante principal de su capacidad
para el cambio evolutivo. Puede mostrarse por experimentos de selección
artificial que las poblaciones naturales albergan un amplio espectro de variaciones
genéticas. La amplitud de la variabilidad genética puede ser cuantificada
comparando las estructuras de las proteínas y, más recientemente,
mediante la secuenciación de las moléculas de DNA .
El equilibrio de Hardy-Weinberg
describe el estado estacionario de las frecuencias alélicas y genotípicas
que existiría en una población ideal en la cual se cumplieran
cinco condiciones. El equilibrio de Hardy-Weinberg demuestra que la recombinación
genética que resulta de la meiosis y de la fecundación no cambia
por sí misma la frecuencia de los alelos en el reservorio génico.
La expresión matemática del equilibrio de Hardy-Weinberg suministra
un método cuantitativo para determinar la intensidad y la dirección
del cambio en las frecuencias alélicas y genotípicas.
El principal factor de
cambio en la composición del reservorio génico es la selección
natural, aunque existen otros procesos involucrados. Estos procesos incluyen
la mutación, el flujo de genes , la deriva genética y el apareamiento
no aleatorio o preferencial.
La reproducción
sexual es el factor más importante que promueve la variabilidad genética
en las poblaciones. Existen otros mecanismos que garantizan la exogamia y que
también contribuyen al incremento de la variabilidad.
Los biólogos evolutivos
proponen que los genes estructurales existentes en la actualidad tuvieron sus
comienzos en muy pocos protogenes. Estos protogenes luego se habrían
duplicado y modificado por la acumulación de mutaciones durante los últimos
4.000 millones de años.
La amplitud de la variabilidad
El parecido evidente que
existe entre los progenitores y sus descendientes se explica por la notable
precisión con la cual el DNA se replica y se transmite de una célula
a sus células hijas durante la división celular.
El DNA de las células
de cualquier individuo es, excepto en el caso de mutaciones ocasionales, una
réplica del DNA que el individuo recibió de sus progenitores.
De hecho, los mecanismos de replicación y transmisión del DNA
no sólo nos vinculan con nuestros antecesores inmediatos, sino que también
expresan la relación que existe entre nosotros y todos los demás
seres vivos.
Aunque la fidelidad de
la duplicación es esencial para la supervivencia de los organismos individuales
que componen una población, para que ocurra evolución deben producirse
variaciones entre los individuos. Estas variaciones constituyen la materia prima
sobre la cual operan las fuerzas evolutivas y son las que hacen posible que
poblaciones sometidas a condiciones diferentes sean diferentes.
Así, la variabilidad es una característica de la población; no existe un tipo ideal sino una gama de variantes que va cambiando en el tiempo y en el espacio.
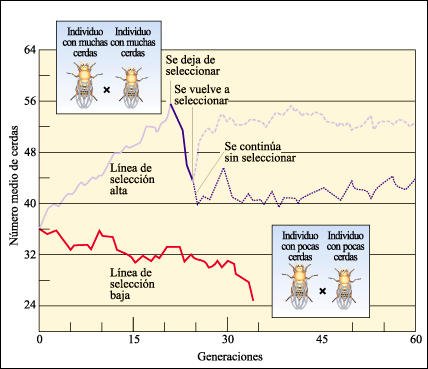
Los resultados de experimentos realizados con Drosophila melanogaster demuestran el grado de variabilidad latente en una población.
En los experimentos realizados con Drosophila, de un único linaje parental,
se seleccionó un grupo para incrementar el número de cerdas de
la superficie ventral (línea de selección alta) y otro para disminuir
el número de cerdas (línea de selección baja). Como puede
verse, la línea de selección alta alcanzó rápidamente
un pico de 56 cerdas pero, luego, el conjunto comenzó a volverse estéril.
Se interrumpió la selección en la generación 21 y se reanudó
en la generación 24. En esta ocasión, se recuperó el alto
número de cerdas anterior y no hubo una pérdida aparente en la
capacidad de reproducción. Nótese que, después de la generación
24, la línea que se reproducía sin selección también
fue proseguida, según lo indica el trazo azul punteado. Después
de 60 generaciones, los miembros del grupo de cruzamientos libres de la línea
de selección alta tuvieron un promedio de 45 cerdas. La línea
de selección baja se extinguió debido a la esterilidad.
La genética de poblaciones
moderna ha indagado de varias maneras diferentes la amplitud de esta variabilidad
y cómo estas variaciones se mantienen en los reservorios génicos.
La selección artificial,
proceso considerado como una analogía directa de la selección
natural, mostró que existe una enorme cantidad de variabilidad oculta
en el reservorio génico, y que esta variabilidad latente puede expresarse
bajo las presiones de la selección.
El análisis a nivel molecular
constituye un método más reciente para estimar la variabilidad.
Sobre la base de que las secuencias de aminoácidos de las proteínas
reflejan las secuencias de nucleótidos de los genes que las codifican
se analizan las proteínas presentes en poblaciones naturales. Se identifican
entonces proteínas (enzimas) funcionalmente diferentes que están
codificadas en diferentes loci. Sobre la base de estos datos, sin analizar directamente
ninguno de los genes, se puede saber cuántos alelos de cada uno de los
genes responsables de codificar la información para cada una de las enzimas
existe en la población y estimar así la variabilidad.
Recientemente, comenzó a ser posible realizar un nivel de análisis que los genetistas evolutivos estaban esperando con ansiedad: el estudio de la variabilidad a nivel último, es decir, a nivel del DNA. Dado que no todos los cambios en los nucleótidos dan como resultado cambios en la secuencia de aminoácidos, y que no todos los cambios en la secuencia de aminoácidos son detectables por electroforesis, se esperaba que el estudio del DNA revelara una mayor variabilidad. Estos estudios se hicieron factibles gracias a la incorporación de algunas técnicas de la biología molecular, tales como la reacción en cadena de la polimerasa -o PCR- que permiten obtener grandes cantidades de DNA a partir de unas pocas moléculas de cadena simple. La posterior secuenciación de estos productos de amplificación han permitido poner de manifiesto un amplio intervalo de variabilidad oculta, representado por mutaciones silenciosas. Estas mutaciones, previamente indetectables, son cambios puntuales en regiones no codificantes o cambios de bases que no modifican el aminoácido codificado y que, por lo tanto, no alteran la proteína resultante.
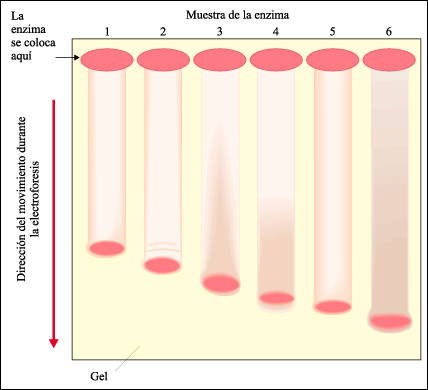
El método que usaron Hubby y Lewontin para analizar las enzimas de Drosophila fue la electroforesis.
En la técnica de electroforesis,
la muestra se disuelve y se coloca en un extremo de una lámina de un
gel al que se le aplica un campo eléctrico débil. La velocidad
con que se mueven las moléculas en este campo eléctrico está
determinada por su tamaño y por su carga eléctrica. Como resultado,
es posible separar las proteínas que tienen diferencias estructurales,
aunque sean muy leves. Este esquema muestra el aspecto de una electroforesis
de seis formas diferentes de una enzima (alozimas). El material de cada columna
fue obtenido de moscas homocigóticas para uno de los seis alelos diferentes
que codifican para la enzima.
Un estado estacionario: el equilibrio
de Hardy-Weinberg
A principios del siglo XX, los genetistas comenzaban a comprender las leyes de la herencia y el origen de nueva variabilidad a partir de la mutación.
Sin embargo, dado que la evolución
es un proceso que se desarrolla a través del tiempo, era necesario indagar
cómo se comportaba la variabilidad presente en una población a
través de las generaciones. Si en una población existen, por ejemplo,
dos alelos para una misma característica que están presentes en
una determinada proporción y en ciertas combinaciones genotípicas,
¿se modificará esta proporción en la siguiente generación,
luego del proceso de reproducción sexual?
Esta pregunta fue respondida,
en 1908, por G. H. Hardy, un matemático inglés, y por G. Weinberg,
un médico alemán. Trabajando de manera independiente, Hardy y
Weinberg mostraron que las combinaciones que resultan del proceso de apareamiento
y reproducción que ocurre en cada generación en los organismos
diploides no involucran un cambio en la composición general del reservorio
génico. Para demostrar esto, propusieron un modelo teórico que
permite examinar el comportamiento de los alelos en una po-blación ideal
en la cual rigen cinco condiciones: 1) No ocurren mutaciones; 2) no hay desplazamiento
neto de individuos con sus genes hacia el interior de la población (inmigración)
o hacia afuera (emigración); 3) la población es lo suficientemente
grande como para que se apliquen las leyes de la probabilidad; o sea, es altamente
improbable que el azar, por sí mismo, pueda alterar la frecuencia de
los alelos; 4) el apareamiento entre individuos es al azar y 5) no hay diferencia
en el éxito reproductivo de los genotipos considerados, es decir, que
el llevar diferentes combinaciones alélicas no confiere ventaja a sus
portadores. La progenie de todos los apareamientos posibles tiene la misma probabilidad
de sobrevivir y reproducirse en la generación siguiente.
Si se considera un único gen con sólo dos alelos, A y a, se puede demostrar matemáticamente que si se cumplen las cinco condiciones mencionadas previamente, entonces las frecuencias, o proporciones relativas, de los alelos A y a en la población no cambiarán de una generación a otra. Más aun, la frecuencia de los tres genotipos posibles de estos alelos -los genotipos AA, Aa y aa- no cambiarán de una generación a la si-guiente. El reservorio génico estará en un estado estacionario -en un equilibrio- con respecto a estos alelos. Así, la ecuación de Hardy-Weinberg establece que en una población ideal, en la cual se cumplan las cinco condiciones planteadas por el modelo, ni las frecuencias alélicas ni las frecuencias genotípicas cambian de una generación a otra.
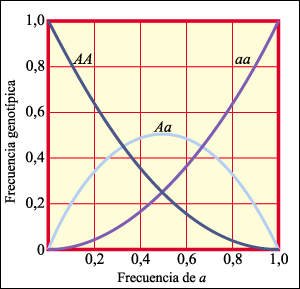
La relación entre la frecuencia del alelo a en la población y la frecuencia de los genotipos AA, Aa y aa.
Naturalmente, cuantos más
AA haya, menor será la frecuencia de a. Dadas las interrelaciones de
los alelos en los genotipos: AA, Aa y aa, un cambio en la frecuencia de uno
u otro alelo da como resultado un cambio correspondiente, y simétrico,
en las frecuencias del otro alelo y de los genotipos.
Considérese un único
gen que tiene sólo dos alelos, A y a. Hardy y Weinberg demostraron matemáticamente
que si se cumplen las cinco condiciones mencionadas previamente, entonces las
frecuencias, o proporciones relativas, de los alelos A y a en la población
no cambiarán de una generación a otra. Más aun, la frecuencia
de los tres genotipos posibles de estos alelos -los genotipos AA, Aa y aa- no
cambiarán de una generación a la siguiente. El reservorio génico
estará en un estado estacionario -en un equilibrio- con respecto a estos
alelos. Este equilibrio se expresa con la siguiente ecuación:
p2
+ 2pq + q2= 1
En esta ecuación, la
letra p designa la frecuencia de un alelo y la letra q designa la frecuencia
del otro, la suma de p y q siempre debe ser igual a l (o sea, p + q representa
el 100% de los alelos de ese gen particular en el reservorio génico).
La expresión p2 designa la frecuencia de individuos
homocigóticos para un alelo, q2 es la frecuencia
de individuos homocigóticos para el otro alelo, y 2pq es la frecuencia
de heterocigotos.
Derivación de la ecuación
de Hardy-Weinberg
Para comprender de qué
manera Hardy y Weinberg obtuvieron su ecuación y demostraron el equilibrio
de las frecuencias alélicas y genotípicas en una población
que cumplía con las cinco condiciones establecidas, observemos más
de cerca el comportamiento del gen con sólo dos alelos, A y a. Nos interesan
las frecuencias relativas -o sea, las proporciones- de A (p) y a (q) de una
generación a la siguiente. Como notamos arriba, cuando hay sólo
dos alelos, la suma de p y q debe igualar a la unidad: p + q = 1.
Por ejemplo, supongamos que,
en una población particular, el 80% de los alelos del gen en estudio
es del tipo A. La frecuencia de A es 0,8, o p = 0,8. Dado que hay sólo
dos alelos, sabemos entonces que la frecuencia del alelo a es 0,2 (q = 1 - p).
Supongamos que las frecuencias
relativas de A y a son iguales tanto en los machos como en las hembras (como
ocurre con la mayoría de los alelos en las poblaciones naturales). Supongamos
ahora que los machos y las hembras se aparean al azar con respecto al hecho
de ser portadores de los alelos A y a. Podemos calcular las frecuencias de los
genotipos resultantes dibujando un tablero de Punnett. Como puede verse en la
figura anterior, las proporciones de los genotipos en la población, producida
por este apareamiento al azar, serán 64% AA, 32% Aa y 4% aa.
En lugar de dibujar un tablero
de Punnett, podemos hacer lo mismo de manera algebraica. Dado que p + q= 1,
se sigue que:
(p+ q) x (p+
q) = 1 x 1 = 1
o, como se recordará
probablemente del álgebra:
p2+
2pq + q2 = 1
Esta expresión algebraica
de las frecuencias genotípicas es la ecuación de Hardy-Weinberg.
Apliquemos esta explicación
al apareamiento al azar que había ocurrido en nuestra población.
Tomando los valores iniciales para la frecuencia de los dos alelos, obtenemos
los siguientes resultados:
p2 =
0,8 x 0,8 = 0,64 (la frecuencia de los genotipos AA)
2 pq= 2 x 0,8 x 0,2 = 0,32 (la
frecuencia de los genotipos Aa)
q2 =
0,2 x 0,2 = 0,04 (la frecuencia de los genotipos aa)
¿Qué ha ocurrido
con la frecuencia de los dos alelos en el reservorio génico como resultado
de esta ronda de apareamientos? Sabemos, por nuestros cálculos, que la
frecuencia AA es 0,64. Además, la mitad de los alelos en los heterocigotos
Aa son A, de modo que la frecuencia total del alelo A es 0,64 más la
mitad de 0,32 (o sea, 0,64 más 0,16), lo cual totaliza 0,8. La frecuencia
(p) del alelo A no ha cambiado. De modo semejante,la
frecuencia total del alelo a es 0,04 (en los homocigotos) más 0,l6 (la
mitad de los alelos en los heterocigotos) o sea 0,2. La frecuencia (q) del alelo
a también ha permanecido constante.
Si ocurre otra ronda de apareamientos,
la proporción de genotipos AA, Aa y aa en nuestra población será
nuevamente de 64%, 32% y 4%, respectivamente. Nuevamente, la frecuencia del
alelo A será 0,8, y la del alelo a 0,2. Y así sucesivamente, generación
tras generación. En una población ideal, en la cual se cumplan
las cinco condiciones planteadas por el modelo, ni las frecuencias alélicas
ni las frecuencias genotípicas cambian de una generación a otra.
El equilibrio de Hardy-Weinberg
y su formulación matemática han sido un valiosísimo fundamento
para la genética de poblaciones. A primera vista, esto parece difícil
de comprender, dado que es prácticamente imposible que las cinco condiciones
requeridas por el modelo para que el equilibrio se mantenga se cumplan en una
población natural.
Si bien las frecuencias de los
alelos en las poblaciones naturales siempre están cambiando, sin la ecuación
de Hardy-Weinberg no sabríamos cómo detectar el cambio, determinar
su magnitud y dirección, o describrir las fuerzas que lo determinan.
Sin embargo, si podemos identificar el genotipo de los individuos de una población,
podremos estimar las frecuencias génicas y comparar estos datos con el
modelo de Hardy-Weinberg. Si hacemos esto durante varias generaciones, podemos
representar con exactitud en un gráfico los cambios que están
ocurriendo en el reservorio génico y, en función de ello, investigar
las causas.
Los agentes del cambio
De acuerdo con la teoría
evolutiva moderna, la selección natural es la fuerza principal que explica
el cambio en las frecuencias de los alelos. Existen, sin embargo, otros agentes
que pueden cambiar las frecuencias de los alelos en una población. Entre
estos agentes pueden distinguirse principalmente la mutación, el flujo
de genes, la deriva genética y el apareamiento no aleatorio.
De acuerdo con la teoría
evolutiva moderna, la selección natural es la fuerza principal que explica
el cambio en las frecuencias de los alelos. Existen, sin embargo, otros agentes
que pueden cambiar las frecuencias de los alelos en una población. Entre
estos agentes pueden distinguirse principalmente la mutación, el flujo
de genes, la deriva genética y el apareamiento no aleatorio.
Las mutaciones ocurren al azar,
o por casualidad. Esto significa que aunque la tasa de mutaciones puede ser
influida por factores ambientales, las consecuencias de las mutaciones son independientes
de las características del ambiente y, por lo tanto, de su potencialidad
para constituirse en un beneficio o en un perjuicio para el organismo y su progenie.
El flujo de genes -la entrada
o salida de los alelos del reservorio génico - pueden introducir nuevos
alelos o alterar las proporciones de los alelos ya presentes y, frecuentemente,
este proceso tiene el efecto de contrarrestar a la selección natural.
La interrupción de flujo génico por alguna barrera geográfica
es un hecho muy importante en el proceso de formación de especies nuevas.
El equilibrio de Hardy-Weinberg
tiene validez sólo si la población es grande. Este requisito es
necesario porque el equilibrio depende de las leyes de la probabilidad. La deriva
genética es un proceso que ocurre generalmente en poblaciones pequeñas
En las poblaciones pequeñas, ciertos alelos pueden aumentar o disminuir
su frecuencia y, a veces, incluso desaparecer, como resultado del azar.
Los genetistas de poblaciones y otros biólogos evolutivos generalmente concuerdan en que la deriva genética desempeña un papel significativo en la determinación del curso evolutivo de las poblaciones. Sin embargo, su importancia relativa, comparada con la de la selección natural, es un asunto que se debate actualmente. Hay, por lo menos, dos situaciones en las cuales se ha demostrado su importancia. Una de ellas es el efecto fundador.
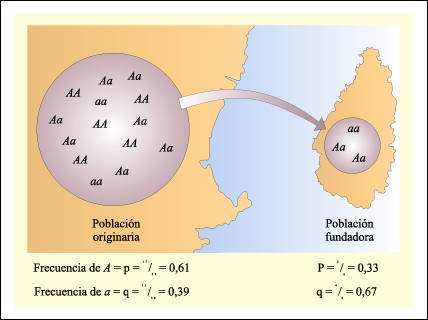
Efecto fundador.
El efecto fundador puede manifestarse
cuando una nueva población es fundada a partir de una pequeña
muestra de una población original (por ejemplo la colonización
de una isla no habitada anteriormente, a partir de unos pocos individuos de
una población continental), las frecuencias alélicas en el grupo
fundador pueden ser diferentes de las presentes en la población de donde
proceden. Como consecuencia de ello, el reservorio génico de la nueva
población tendrá una composición diferente al reservorio
de la población originaria.
Otro caso de deriva genética
aparece cuando el número de miembros de una población se reduce
drásticamente por un acontecimiento que tiene poca o ninguna relación
con las presiones habituales de la selección natural. A este fenómeno
se lo denomina cuello de botella.
El apareamiento no aleatorio
o preferencial provoca cambios en las proporciones de los genotipos y puede
o no afectar las frecuencias alélicas. Una forma de apareamiento no aleatorio,
particularmente importante en las plantas, es la autopolinización. En
los animales, el apareamiento no aleatorio depende, a menudo, del comportamiento.
Este apareamiento no aleatorio es un componente importante de selección
natural en algunas especies. El apareamiento no aleatorio puede provocar cambios
en las frecuencias genotípicas sin producir necesariamente ningún
cambio en la frecuencia de los alelos en cuestión.
Preservación y promoción
de la variabilidad
Sin duda, el mecanismo más
importante por el cual se promueve la variabilidad de la progenie en los organismos
eucarióticos es la reproducción sexual y lo hace de tres modos:
1) por distribución independiente de los cromosomas en la meiosis §;
2) por crossing-over con recombinación genética en la meiosis
y 3) por la combinación de los dos genomas parentales en la fecundación
.
En cada generación, los
alelos son distribuidos en combinaciones nuevas. En contraste con esto, los
organismos que se reproducen sólo asexualmente mediante procesos en los
que intervienen la mitosis y la citocinesis, pero no la meiosis -excepto en
el caso de que haya ocurrido una mutación durante el proceso de duplicación-
el organismo nuevo será exactamente igual a su único progenitor.
Con el tiempo se formarán muchos clones; cada uno de los cuales podrá
llevar una o más mutaciones pero, a menos que las mismas mutaciones ocurran
en los mismos clones, las combinaciones potencialmente favorables nunca se acumularán
en un mismo genotipo.
En cuanto a las desventajas,
los organismos que se reproducen sexualmente sólo pueden hacerlo a la
mitad de la velocidad que los organismos que se reproducen asexualmente. La
única ventaja para el organismo que se reproduce sexualmente es la promoción
de la variabilidad, la producción de nuevas combinaciones de alelos entre
la progenie. Por qué esta variabilidad resulta ventajosa para el organismo
individual es objeto de una antigua y larga discusión que aún
sigue abierta
En las poblaciones que se reproducen
sexualmente se han desarrollado muchos mecanismos que promueven nuevas combinaciones
genéticas.
Estos mecanismos incluyen la presencia de alelos de autoesterilidad y de adaptaciones anatómicas que inhiben la autofecundación en las plantas y de diversas estrategias del comportamiento que inhiben la cruza entre organismos emparentados, en los animales. La variabilidad es también preservada por la diploidía, que protege a los alelos recesivos raros de la selección natural. La selección natural también puede promover y preservar la variabilidad. En los casos de superioridad de los heterocigotos, por ejemplo, se selecciona al heterocigoto con preferencia a cualquier homocigoto, manteniendo así a ambos alelos en la población. La heterosis, o vigor híbrido, es el resultado de la superioridad del heterocigoto o bien del enmascaramiento en heterocigosis de los posibles efectos perjudicales de alelos recesivos.
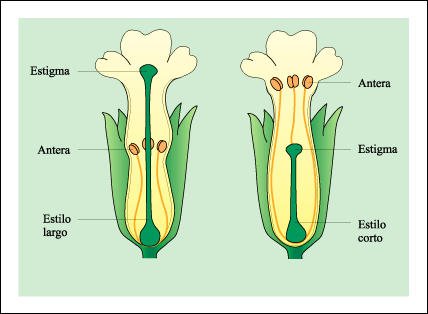
Dibujos de dos tipos de flores de la misma especie que representan un ejemplo de heterostilia (distinta longitud de los estilos).
En la figura anterior puede notarse que las anteras con polen de la flor con
estilo largo y el estigma de la flor con estilo corto, se encuentran a la misma
altura, en medio del tubo floral. Al mismo tiempo, el estigma de la flor con
estilo largo coincide con las anteras de la flor de estilo corto. Un insecto
que visitara estas flores en busca de néctar, recogería polen
en áreas diferentes de su cuerpo, de modo tal que el polen de la flor
con estilo corto probablemente será depositado sobre los estigmas de
la flor con estilo largo, y viceversa.
El origen de la variabilidad
genética
Las nuevas técnicas de
análisis del DNA de los cromosomas de los organismos eucarióticos
ha permitido comprobar que grandes segmentos de DNA -los transposones - tienen
la capacidad para producir duplicados de sí mismos y dispersar estos
duplicados en otros sitios del mismo cromosoma o de otros cromosomas. Estos
genes duplicados son entonces libres para transitar su propio camino evolutivo,
dejando que sus funciones sean desempeñadas por los genes parentales
originales. Los genes duplicados están libres, por lo tanto, de restricciones
selectivas, permitiendo que se acumulen las mutaciones.
Los biólogos evolutivos
proponen que los genes estructurales existentes actualmente tuvieron sus comienzos
en muy pocos protogenes, que luego se duplicaron y modificaron por la acumulación
de mutaciones durante los últimos 4.000 millones de años. Más
importante aun es que existen evidencias claras de que este proceso de duplicación
y subsiguiente mutación continúa en el presente. La duplicación
y la modificación génica han desempeñado indudablemente
un papel muy importante en la evolución. Es probable que, a medida que
se incremente nuestra comprensión acerca de estos procesos, se requerirá
una revisión de algunos aspectos de la teoría evolutiva.


