Capítulo 22.
La selección natural
De acuerdo con el propio relato de Darwin, el
concepto de selección natural se le ocurrió en 1838 leyendo el
"Ensayo sobre el principio de población" de Malthus. Darwin
comprendió que todas las poblaciones -no sólo la población
humana- están condenadas potencialmente a exceder los recursos de los
que depende su existencia. Sólo una pequeña fracción de
los individuos que podrían existir, nace, sobrevive y llega a reproducirse.
Según Darwin, los que sobreviven son los que se encuentran "favorecidos",
para usar su propio término, por ser portadores de ligeras variaciones
ventajosas. Este proceso de mayor supervivencia y reproducción de los
"favorecidos" fue llamado por él selección natural,
por analogía con la selección artificial practicada por los criadores
de animales y plantas domésticos.
La selección natural
se define como la reproducción diferencial de genotipos que resulta de
las interacciones entre los organismos individuales y su ambiente y, de acuerdo
con la Teoría Sintética de la evolución, es la principal
fuerza de la evolución. La selección natural puede actuar produciendo
cambios o manteniendo la variabilidad dentro de una población.
La selección natural
puede operar solamente sobre las características expresadas en el fenotipo.
La unidad de selección es el fenotipo completo: la totalidad del organismo.
En casos extremos, un sólo alelo puede ser decisivo en la selección
pero, generalmente, un fenotipo exitoso es el resultado de la interacción
de muchos genes.
Las tres tipos principales de selección natural son la selección
normalizadora, la selección disruptiva y la selección direccional.
Otro tipo de selección es la selección dependiente de lafrecuencia
y una quinta categoría es la selección sexual.
La selección natural
implica interacciones entre organismos individuales, su ambiente físico
y su ambiente biológico -es decir, con otros organismos-. Generalmente,
el resultado de la selección natural es la adaptación -aunque
imperfecta- de las poblaciones a su ambiente. La adaptación al ambiente
biológico resulta de la interacción recíproca de especies
de organismos, es decir, de la coevolución . La postura clásica
que considera a la evolución como un proceso de creciente adaptación
a partir de la acción de la selección natural ha recibido numerosas
críticas.
Se han propuesto que la
fijación azarosa de rasgos neutrales, los procesos de alometría
y heterocronía, el efecto pleiotrópico y el ligamiento genético,
los emergentes arquitectónicos y las variaciones ambientales sin base
genética pueden dar origen a nuevas características, en forma
alternativa a la selección natural. Recientemente, se han introducido
nuevos conceptos relativos a la adaptación para distinguir la función
actual de una estructura del proceso que explica su origen. Entre ellos, se
cuentan la exaptación, la no aptación y la aptación, para
denominar conjuntamente a las exaptaciones y las adaptaciones.
Selección natural
y mantenimiento de la variabilidad
En el curso de las controversias
que llevaron a la formulación de la Teoría Sintética, algunos
biólogos argumentaron que la selección natural serviría
sólo para eliminar al "menos apto" y, en consecuencia, tendería
a reducir la variación genética de las poblaciones, actuando en
este sentido como una fuerza antievolutiva. La genética de poblaciones
moderna ha demostrado que esto no es cierto. La selección natural puede
ser un factor crítico para preservar y promover la variabilidad en una
población.
Hay muchos ejemplos de
cómo puede mantenerse la variabilidad en los que diversas fuerzas selectivas
puedan estar operando simultáneamente. Un buen ejemplo lo constituyen
el color y los patrones de bandeado en caracoles.
En distintas especies
de caracoles terrestres del género Cepaea coexisten diversas coloraciones
de la concha del caracol. Además, la concha puede presentar hasta cinco
bandas longitudinales de color. La evidencia fósil indica que estos diferentes
tipos de conchas han coexistido durante más de 10.000 años. Los
caracoles presentan un tipo de polimorfismo denominado polimorfismo equilibrado,
en contraposición al polimorfismo llamado transitorio. En los ambientes
uniformes, por ejemplo, hay una frecuencia más alta de caracoles sin
bandas, mientras que en los hábitat irregulares y variados, como los
pisos de los bosques, la mayoría tiende a ser bandeada. De modo análogo,
los hábitat más verdes tienen la mayor proporción de conchas
amarillas, pero entre los caracoles que viven sobre fondos oscuros, las conchas
amarillas son mucho más visibles, resultando claramente desventajosas
a juzgar por el éxito de captura de este tipo de concha por los zorzales,
sus predadores naturales. Sin embargo, se encuentran ambos tipos de conchas
en las distintas colonias, distantes unas de otras.
Parecen existir factores
fisiológicos que están correlacionados con los diferentes patrones
de coloración de las conchas, dado que los genes que controlan ambos
aspectos conformarían un grupo de ligamiento. Esto explicaría,
aunque no de manera concluyente, por qué están presentes los dos
tipos. Se han hecho experimentos que muestran, por ejemplo, que los caracoles
sin bandas (especialmente los amarillos) son más resistentes al calor
y al frío que aquellos que presentan bandas. En otras palabras, es probable
que el polimorfismo se haya mantenido porque están operando varias presiones
selectivas diferentes, las cuales actúan en forma conjunta.
El resultado de estas
interacciones es el mantenimiento de las variaciones genéticas que determinan
el color y la formación de bandas, de modo que se establece un polimorfismo
equilibrado.
Tipos de selección
Las tres categorías
principales de la selección natural son: la selección normalizadora,
en la cual se eliminan los fenotipos extremos de la población; la selección
disruptiva, en la que se seleccionan los fenotipos extremos a expensas de formas
intermedias; y la selección direccional, en la que uno de los extremos
es favorecido, empujando a la población a lo largo de una vía
evolutiva particular.
Otro tipo de selección
es la selección dependiente de la frecuencia, en la cual la aptitud de
un fenotipo disminuye a medida que se hace más común en la población
y se incrementa a medida que se hace menos frecuente. Una quinta categoría,
la selección sexual, es el resultado de la competencia en la búsqueda
de pareja; puede aumentar en gran medida la reproducción diferencial,
sin mejorar la adaptación a otros factores ambientales.
El resultado de la seleccion
natural: la adaptación
La selección natural
da como resultado la adaptación, con sus varios significados y manifestaciones
múltiples. Implica interacciones entre organismos individuales, su ambiente
físico y su ambiente biológico -es decir, con otros organismos-.
En muchos casos, las adaptaciones que resultan de la selección natural
pueden correlacionarse claramente con factores ambientales o con las presiones
selectivas ejercidas por otros organismos.
Algunas variaciones fenotípicas
intraespecíficas siguen una distribución geográfica y pueden
ser correlacionadas con cambios graduales de temperatura, humedad o alguna otra
condición ambiental. Esta variación gradual de una característica
o de un complejo de características en correlación con un gradiente
ambiental es conocida como clina.
Por otra parte, una especie
que ocupa muchos hábitat diferentes puede presentar características
ligeramente diferentes en cada uno de ellos. Cada uno de estos grupos que presenta
fenotipos diferentes es conocido como un ecotipo. Estas diferencias entre los
ecotipos ¿están determinadas completamente por el ambiente?, ¿o
representan adaptaciones resultantes de la acción de la selección
natural sobre la variación genética?
Experimentos realizados
en la planta P. glandulosa demostraron que muchas de las diferencias fenotípicas
halladas entre los ecotipos de P. glandulosa se debían a diferencias
genéticas. No resulta sorprendente que en ambientes muy diferentes se
hayan seleccionado distintas características. A lo largo del tiempo,
las diferencias genéticas entre las plantas individuales han terminado
por expresarse como diferencias genéticas entre los subgrupos de la población
de P. glandulosa, que constituyen los actuales ecotipos. Este proceso puede
ser el primer paso en la formación de nuevas especies.
Cuando las poblaciones
de dos o más especies establecen interacciones tan estrechas que cada
una ejerce una notable fuerza selectiva sobre la otra, ocurren ajustes simultáneos
que dan como resultado un proceso de coevolución. Uno de los más
importantes, en términos del número de especies e individuos que
intervienen, es la coevolución de las flores y sus polinizadores o el
de las a plantas e insectos, aquellos dos aliados y enemigos ancestrales.
La postura clásica
que considera a la evolución como un proceso de creciente adaptación
a partir de la acción de la selección natural ha recibido numerosas
críticas. Darwin expresó su cautela acerca del alcance explicativo
de la selección natural, cuando en el prólogo de El origen de
las especies afirmaba: "...estoy convencido de que la selección
natural ha sido el medio más importante, si bien no el único,
de modificación". Sin embargo, a partir de 1940, los genetistas
que adherían a la Teoría Sintética endurecieron su postura
alrededor de este argumento.
En el marco de la síntesis
evolutiva, toda característica de un organismo era interpretada como
una adaptación y, por lo tanto, como el resultado del proceso de selección
natural. Dos décadas más tarde, se comenzó abandonar esta
ortodoxia.
Por una parte, la teoría
neutralista plantea que la mayoría de las variantes genéticas
a nivel molecular no confieren ventaja ni desventaja al portador, por lo que
se fijan o se pierden por deriva genética. Por otra parte se ha cuestionado
la idea de que la selección natural es capaz de producir adaptaciones
óptimas, ya que las posibilidades de cambio están drásticamente
limitadas tanto por factores intrínsecos -como el programa genético,
los patrones de desarrollo y la estructura del organismo-, como por factores
extrínsecos -como la constante modificación del nicho que hace
que la especie siempre esté un paso atrás de los requerimientos
del ambiente-.
Otra de las críticas
planteadas apunta a cuestionar la omnipotencia de la selección natural,
es decir, ¿puede la selección natural, actuando en forma constante,
perfeccionar crecientemente las características de las especies hasta
llegar a producir adaptaciones óptimas? La respuesta de los críticos
es un no rotundo, ya que la selección natural no hace "lo que quiere"
sino "lo que puede". Si la selección natural, operando a través
del cambio en las frecuencias génicas, fuera el único proceso
capaz de explicar la diversidad de la vida, las posibilidades serían
ilimitadas.
Si bien toda adaptación es una característica organísmica que se establece gradualmente, mediante un proceso de selección natural que permite acumular las pequeñas variaciones favorables, es necesario comprender que no toda característica de un organismo representa una adaptación. Los biólogos evolutivos que cuestionan esta postura la denominan panseleccionista.
El diseño en forma de bóveda en abanico de este techo implica que necesariamente debe existir una estructura central formada por dos triángulos enfrentados. Parecen importantes pero, ¿tiene acaso una finalidad?
Estos biólogos no niegan
la existencia de la adaptación, pero desplazan el interés a una
mirada más plural, que permite considerar al organismo globalmente y
recurrir a explicaciones alternativas a la selección natural para interpretar
el origen de sus características. Conforme a los diferentes procesos
que operan en las poblaciones, es posible comprender que algunos caracteres
se fijan por deriva genética, es decir que su presencia se debe simplemente
al azar. También es necesario considerar que una parte de la variación
representa un ajuste al medio sin base genética, de modo que no se ha
establecido mediante selección natural.
Se ha propuesto que en muchos casos las nuevas características pueden surgir mediante mecanismos alternativos a la selección natural tales como la fijación azarosa de rasgos neutrales, los procesos de alometría y heterocronía, el efecto de la pleiotropía y el ligamiento genético, los emergentes arquitectónicos y las variaciones ambientales sin base genética.
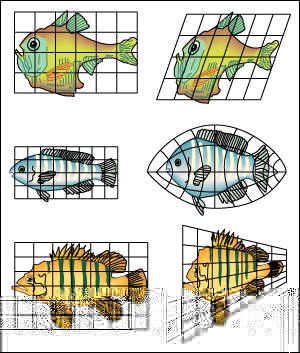
Una simple transformación, dada por el crecimiento diferencial de distintas partes de un mismo plan estructural (alometrías), podría explicar diferencias complejas entre especies.
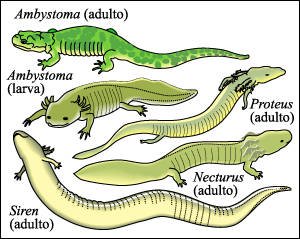
Los cambios de los tiempos de desarrollo (heterocronías) pueden producir novedades evolutivas.
Las salamandras adultas de los géneros Proteus, Necturus y Siren tienen
branquias toda su vida. Esta es una característica neoténica si
se las compara con las etapas correspondientes a larva y adulto de algunas de
las especies de salamandras del género Ambystoma.
Por otra parte, se ha discutido
que no siempre la función que desempeña una estructura en un tiempo
dado revela el proceso que explica su establecimiento. En muchos casos, estructuras
que se han establecido por procesos alternativos a la selección natural
pasan a cumplir una función adaptativa y son modeladas por la selección
natural.
En otros casos, estructuras
que se establecieron gradualmente por selección natural desarrollando
una cierta función, pasan a desempeñar una función diferente
en otra etapa de la evolución. Para estos casos se ha introducido el
concepto de exaptación para denominar a las características que
son incorporadas selectivamente a partir de otra previamente existente, el concepto
de no aptación para denominar a las características neutrales
y el de aptación para denominar conjuntamente a las exaptaciones y las
adaptaciones. Esta terminología permite distinguir la función
actual de una estructura del proceso que explica su origen.


