Capítulo 25.
El comportamiento animal
Las características de
comportamiento de los organismos son producto de la selección natural
y afectan su aptitud . El estudio del comportamiento y su evolución es
un área muy activa en la investigación biológica, que involucra
a científicos de una gran variedad de disciplinas. La etología
es el estudio del comportamiento de los animales en su ambiente natural, con
énfasis particular en los patrones de comportamiento específicos
de especie y sus orígenes evolutivos.
Aunque todo comportamiento tiene
sus raíces en el programa genético de cada individuo, sólo
en pocos casos ha sido posible correlacionar un comportamiento particular con
la presencia o la ausencia de alelos específicos. Los comportamientos
son típicamente el resultado de las interacciones de un gran número
de genes, cuya influencia es ulteriormente modificada por las interacciones
del individuo con su ambiente.
Una explicación completa
acerca de por qué un animal tiene un comportamiento particular implica
diversos tipos de preguntas: preguntas en términos de la secuencia inmediata
de los hechos fisiológicos que llevaron al comportamiento observado y
preguntas acerca de cuál es el valor adaptativo de ese comportamiento,
así como cuáles son sus orígenes evolutivos.
Los patrones de comportamiento
que aparecen esencialmente completos la primera vez que el organismo se encuentra
con el estímulo pertinente se conocen como patrones de acción
fija. Los patrones de acción fija son sumamente estereotipados y rígidos
y, para miembros de un especie dada de edad, sexo y condición fisiológica
particular, son tan predecibles y constantes como las características
anatómicas de la especie.
El aprendizaje es la modificación
del comportamiento como resultado de la experiencia. Se pueden reconocer varias
categorías de aprendizaje, entre ellas, la habituación , el aprendizaje
asociativo -que incluye tanto el condicionamiento clásico como al condicionamiento
operativo-, el troquelado y la imitación.
La gran mayoría de los
organismos obtienen información del ambiente (incluidos otros organismos)
a través de sus sentidos y, con esa información, pueden responder
a los cambios que se van presentando. La comunicación entre dos organismos
lleva implícita la transmisión de información que consiste
en la interacción entre dos organismos. La información se transmite
a través de un canal e involucra la transmisión de una señal.
Los animales se comunican por medio de los más diversos estímulos:
químicos, lumínicos, sonoros, eléctricos, vibratorios,
de presión o de temperatura. Muchos animales viven en sociedades -grupos
de individuos de la misma especie que conviven de una manera organizada, con
división de los recursos y del trabajo y con dependencia mutua-. Desde
un punto de vista evolutivo, es muy interesante el denominado comportamiento
social de los animales. Este comportamiento puede ser "egoísta",
"cooperativo", "altruista" o "malicioso", con
diferentes consecuencias para la aptitud del dador y el receptor.
Las sociedades de insectos se
cuentan entre las más grandes y más complejas de las sociedades
animales.
El comportamiento y la fisiología
de los miembros de la colmena son controlados por el intercambio de señales
químicas en forma de feromonas.Las sociedades de vertebrados frecuentemente
están organizadas en términos de dominancia social. Las jerarquías
de dominancia social pueden tomar distintas formas. La territorialidad es un
sistema de dominancia, por el cual sólo los animales con territorio se
reproducen y los animales con mejores territorio se reproducen más.
La hipótesis de la selección
por parentesco -que introduce el concepto de aptitud inclusiva- presenta una
nueva explicación evolutiva para muchos patrones de comportamiento social.
Los conceptos previos de aptitud evolutiva se habían enfocado sobre el
número relativo de progenie sobreviviente en generaciones futuras. La
aptitud inclusiva se concentra en el número relativo de alelos del individuo
que pasan de una generación a la siguiente como resultante del éxito
reproductivo -ya sea del individuo o de individuos emparentados-.
Desde el punto de vista de la
aptitud inclusiva, algunos científicos sostienen que el comportamiento
social está regulado por el "gen egoísta", que programa
al individuo no necesariamente para el bienestar del individuo ni para sobrevivir,
sino para la perpetuación del alelo por cualquier media. La competencia
por la representación en el reservorio génico futuro puede ocurrir
no sólo entre individuos que compiten obviamente por los mismos recursos
-como alimentos o pareja-, sino también entre padres y sus hijos y entre
los miembros de una pareja reproductora. El altruismo es un comportamiento que
acarrea un costo para el individuo que lo realiza y beneficia a otro u otros
individuos. Se piensa que algunos actos de altruismo están basados en
la aptitud inclusiva; otros pueden estar basados en el altruismo recíproco,
el desempeño de un acto no egoísta con la expectativa de que será
retribuido.
Resulta muy tentador trazar
paralelismos entre el comportamiento de los humanos y el observado en otras
especies. Hasta qué punto los conceptos concernientes a la evolución
del comportamiento animal pueden extrapolarse a la especie humana es una cuestión
que se debate actualmente. Mientras que algunos biólogos sostienen que
la especie humana básicamente no es diferente de cualquier otra especie
y que su comportamiento está determinado por sus genes, un grupo de opositores
opina que los humanos modernos son también producto de la interacción
de los genes con el ambiente (que incluye la cultura) y que, por consiguiente,
aquellos análisis ya no son válidos. Además, los opositores
recurren a otros argumentos basados en los múltiples factores que afectan
el desarrollo de un individuo. Muchos científicos advierten sobre los
peligros que acarrea el concepto de que la biología determina la conducta
humana, ya que estas ideas yacen en las raíces de todas las nociones
de superioridad racial y han proporcionado la fundamentación para la
esclavitud, la explotación y el genocidio.
La bases del comportamiento
Los productos de determinados
genes actúan, en general, de manera indirecta sobre el comportamiento
final de un individuo, por ejemplo, influyendo en el desarrollo de patrones
en el sistema nervioso que mediarán la respuesta comportamental. Por
otra parte, muchos comportamientos están influidos por un gran número
de genes. Estos aspectos del comportamiento a menudo dificultan el estudio de
su base genética.
Sin embargo, el papel de los
componentes genéticos ha podido ser demostrado en un comportamiento simple:
la limpieza de la colmena por parte de la abeja Apis mellifera. En ocasiones,
las larvas que se desarrollan en las celdas de una colmena se enferman y pueden
morir. En algunas cepas, las abejas obreras abren o destapan las celdas que
contienen larvas enfermas y las eliminan, manteniendo así la limpieza
de la colmena. A estas abejas se las llamó "higiénicas",
en contraste con las de cepas "no higiénicas" que no presentan
este comportamiento.
Estudios realizados por medio
de cruzamientos entre abejas "higiénicas" y " no higiénicas"
revelaron una segregación mendeliana simple de rasgos comportamentales.
Sin embargo, aunque los resultados sugieren la existencia de un gen para determinado
comportamiento ("para destapar las celdas" y otro "para eliminar
las larvas" en las abejas "higiénicas"), es importante
tener presente la complejidad de este comportamiento. En la mayoría de
los comportamientos, los mecanismos genéticos son considerablemente más
complicados; en algunos casos, los genes con efectos múltiples -genes
pleiotrópicos - desempeñan un papel importante.
Las mosquitas de la fruta que
tienen el alelo Hk, por ejemplo, son más activas que las que carecen
de este alelo, saltan violentamente cuando una sombra pasa sobre ellas, exhiben
un comportamiento de apareamiento anormal, y muestran movimientos rápidos
de las patas cuando se las anestesia. Estas moscas tienen un lapso de vida más
corto que las mosquitas sin este alelo. Más frecuentemente, las características
de comportamiento son el resultado de herencia, o sea, dependen de la acción
integrada de los alelos de un gran número de genes.
Las preguntas acerca del comportamiento
Se pueden plantear infinidad
de preguntas acerca de un determinado comportamiento: qué neuronas están
involucradas en una cierta actividad motora, qué receptores sensoriales
son los que reciben el estímulo del medio externo, qué áreas
del cerebro son metabólicamente más activas durante el desarrollo
de una secuencia de aprendizaje y hasta qué genes se activan previo a
la adquisición de un determinado comportamiento. Por otra parte, se puede
preguntar acerca de cuál es la causa evolutiva de ese comportamiento:
cuál es su valor adaptativo, cómo permite al animal evitar, por
ejemplo, la depredación y así sobrevivir hasta la madurez, reproducirse
exitosamente y dejar más descendencia que otros animales de la misma
población que no presentan ese comportamiento.
Sobre esta base, es posible
agrupar las preguntas en dos grandes puntos de vista: preguntas en términos
de la secuencia inmediata de los hechos fisiológicos que llevaron al
comportamiento observado y preguntas acerca de cuál es el valor adaptativo
de ese comportamiento, así como cuáles son sus orígenes
evolutivos.
Patrones de acción fija
Comportamientos como la apertura
de la boca de un pichón, la succión por un bebé, o el lengüetazo
de un sapo intentando capturar una mosca se desarrollan con un mínimo
de experiencia sensorial. En estos casos, el patrón de comportamiento
es innato y aparece esencialmente completo la primera vez que el organismo se
encuentra con el estímulo pertinente.
Estos comportamientos son altamente
estereotipados, rígidos y predecibles y además, no tienen un control
de retroalimentación externo, es decir.
Una vez que un comportamiento
de este tipo se dispara, prosigue hasta su finalización. A este tipo
de comportamiento se lo conoce como patrón de acción fija.
Los patrones de acción
fija son específicos de cada especie. Si se comparan entre individuos
de una especie dada, que poseen la edad, el sexo y la condición fisiológica
adecuados, resultan tan específicos y constantes como las características
anatómicas de ese individuo.
Los patrones de acción
fija son iniciados por estímulos externos conocidos, los estímulos
señal. Existen ciertos mecanismos preestablecidos -los mecanismos de
liberación innatos- por los cuales ciertas áreas específicas
dentro del cerebro responden a los estímulos señal.
Un patrón de acción
fija no se manifiesta hasta que el organismo se encuentra con el estímulo
señal apropiado, que entonces estimula el mecanismo de liberación
innato, poniendo en acción la secuencia de movimientos que constituyen
el comportamiento.
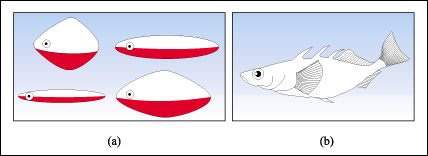
Los colores son la señal que desencadena muchos patrones de acción fija en diferentes especies animales.
Modelos de un pez macho usados por Niko Tinbergen en sus experimentos con el pez espinoso.
a) Modelos groseros de peces machos, pintados de rojo en la superficie ventral.
Estos modelos generaron reacciones mucho más intensas en los machos territoriales
y en las hembras del pez espinoso (agresividad en los machos y atracción
en las hembras) que b), la réplica exacta de un macho sin color.
Además, el organismo
debe encontrarse en un estado fisiológico adecuado para responder; en
otras palabras, debe tener un cierto nivel de motivación. El nivel de
motivación influye en la potencialidad del animal para llevar a cabo
un comportamiento.
Aprendizaje
Todos los patrones de comportamiento,
aun aquellos que parecen relativamente completos al observarse por primera vez,
dependen no sólo de las características del ambiente sino también
del desarrollo fisiológico normal del animal. Un gran número de
comportamientos requiere también de aprendizaje, un proceso en el cual
las respuestas del organismo se modifican como resultado de la experiencia.
Una de las formas más
simples del aprendizaje es la habituación, en la que un organismo reduce
o suprime la respuesta a un estímulo persistente (como el caso de las
palomas de las plazas que dejan de manifestar la reacción de huida frente
a personas y autos). La habituación se diferencia de la fatiga que se
manifiesta como una incapacidad para responder, que revierte con la recuperación,
y de la adaptación sensorial, que consiste en una disminución
temporaria del nivel de respuesta de los receptores frente a un estímulo.
Un tipo de aprendizaje familiar
para todos nosotros es el aprendizaje asociativo en el que un estímulo
llega a conectarse, por medio de la experiencia, con otro estímulo (por
ejemplo, el caso de los perros que se excitan al ver su correa porque la asocian
con el paseo).
Los primeros estudios científicos
de aprendizaje asociativo fueron realizados por I. Pavlov. En su serie de experimentos
sobre condicionamiento clásico, Pavlov mantuvo en cautiverio a un perro
hambriento al que le ofrecía regularmente pequeñas porciones de
alimento. Pavlov logró que el animal estableciera una asociación
entre un estímulo externo -como el sonido de una campana- y el alimento.
Pavlov denominó al alimento estímulo no condicionado, ya que provocaba
la salivación -una respuesta no condicionada- incluso en ausencia de
cualquier otro estímulo. A la señal externa la llamó estímulo
condicionado ya que evocaba una respuesta -la respuesta condicionada- sólo
después de que el perro había sido condicionado a asociar la señal
con la recompensa alimenticia.
El aprendizaje por ensayo y
error, en el que se asocia una actividad particular con un castigo o premio,
se conoce como condicionamiento operante.En
este caso, el animal aprende a asociar su propio comportamiento con las consecuencias
de ese comportamiento a través de la experiencia operativa. Este aprendizaje
implica el establecimiento de memorias, cuya posible localización ha
sido objeto de numerosas investigaciones. En ambientes naturales, el aprendizaje
asociativo frecuentemente implica ensayo y error.
El fenómeno de impronta
o troquelado (en inglés, imprinting) descripto por K. Lorenz implica
aprendizaje asociativo en íntima relación con el desarrollo de
la discriminación.
La discriminación de
los miembros de la propia especie con respecto a los miembros de las otras especies
puede estar basada en una variedad de señales y es de importancia vital
para el éxito reproductivo final de muchos animales. En muchas especies
de aves, este aprendizaje ocurre durante un período crítico específico
en la vida temprana del individuo y depende de la exposición a características
particulares del o los progenitores. Este tipo de aprendizaje es el que constituye
el troquelado.
La imitación de los estímulos
experimentados durante el período crítico es un factor en el aprendizaje
del canto de los pájaros. Éste y otros tipos de aprendizaje imitativo
fueron observados en aves y mamíferos.
El uso de la información:
comunicación
La comunicación entre
dos organismos lleva implícita la transmisión de información
que consiste en la interacción entre dos organismos. La información
se transmite a través de un canal e involucra la transmisión de
una señal.
Los animales se comunican por
medio de los más diversos estímulos: químicos, lumínicos,
sonoros, eléctricos, vibratorios, de presión o de temperatura.
Cada canal presenta ventajas y desventajas que dependen de las condiciones del
ambiente y de las capacidades sensoriales y motoras de cada organismo.

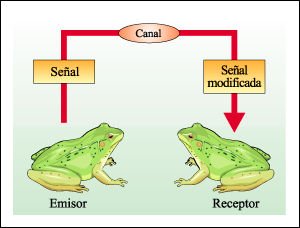
Representación esquemática de la comunicación entre dos organismos.
El emisor emite un mensaje que
se transmite a través de un canal y es percibido, frecuentemente con
modificaciones, por el receptor.
Comportamiento social: introducción
De todos los comportamientos,
tal vez el más enigmático es el de las interacciones que ocurren
entre animales que viven en sociedades estructuradas. Una sociedad es un grupo
de individuos de la misma especie § que viven juntos de manera organizada,
con división de los recursos, del trabajo y en dependencia mutua. Los
estímulos que intercambian y mediante los que se comunican los miembros
del grupo los mantienen juntos y mantienen la estructura social.
Cuando los biólogos evolutivos
comenzaron a analizar el comportamiento social, surgieron algunas cuestiones
perturbadoras como la existencia de castas estériles, o de sociedades
de vertebrados en las que sólo algunos machos procrean. Se definió
entonces el altruismo, un comportamiento que beneficia a otros y es llevado
a cabo con cierto riesgo o costo para quien lo practica. Pero, ¿cómo
se podían explicar los actos de altruismo en términos de la selección
natural que actúa sobre el organismo individual?
Con las subsiguientes observaciones y estudios, se encontró que, obviamente, no todos los comportamientos son altruistas. En 1964, W. D. Hamilton propuso una clasificación de comportamientos sociales.

Adaptado de W.D. Hamilton "La evolución del comportamiento social". Journal of Theoretical Biology, vol 7, p. 1-52.
Cada uno de estos comportamientos
tiene un efecto potencial diferente sobre la aptitud directa (o fitness), o
sea, sobre el éxito reproductivo tanto del individuo que lleva a cabo
el comportamiento -o dador- como del receptor del comportamiento.
Algunos biólogos consideran
que la evolución del comportamiento egoísta a través de
la acción de la selección natural no plantea problemas para la
teoría de la evolución y que es, en realidad, esperable. Aunque
el mantenimiento de la cooperación por selección natural es relativamente
fácil de comprender, el mecanismo por el cual podría haber evolucionado
inicialmente es tan difícil de imaginar como el mecanismo que subyace
a la evolución del altruismo.
Sociedades de insectos
Las sociedades de insectos se
encuentran entre las más antiguas de todas las sociedades y, junto con
las modernas sociedades humanas, están entre las más complejas.
Los insectos sociales incluyen las termitas y los himenópteros (hormigas,
avispas y abejas).
La mayoría de las especies
vivientes de abejas y avispas son solitarias, y otras muestran grados variables
de sociabilidad. Es posible construir un escenario de las distintas etapas de
la evolución social mediante el análisis de las especies que intervienen.
En las especies solitarias,
la hembra construye un pequeño nido, deposita sus huevos en él,
lo provee de reservas alimenticias, y lo abandona para siempre. Habitualmente
muere poco después, de manera que no hay superposición de generaciones.
Entre las especies presociales
o subsociales, la madre regresa para alimentar a las larvas durante cierto tiempo
y la prole producida puede, posteriormente, depositar sus huevos en el mismo
nido o panal. En estos casos, la colonia no es permanente, no hay división
del trabajo y todas las hembras son fértiles.
Los insectos eusociales se caracterizan
por la cooperación en el cuidado de la prole y una división del
trabajo en la que los individuos estériles trabajan en beneficio de los
que se reproducen. Todas las hormigas y las termitas y algunas especies de avispas
y abejas -por ejemplo, las abejas melíferas- son eusociales.
En la colonia de abejas melíferas
(y entre otros insectos eusociales también), la reina es la única
forma femenina reproductora. Ella y su progenie son atendidas por obreras estériles.
Una abeja que retorna de una fuente de alimento se comunica con las abejas que se encuentran dentro de la colmena. Puede realizar dos tipos de danza dentro de la colmena, en completa oscuridad: la danza circular y la danza "en ocho".
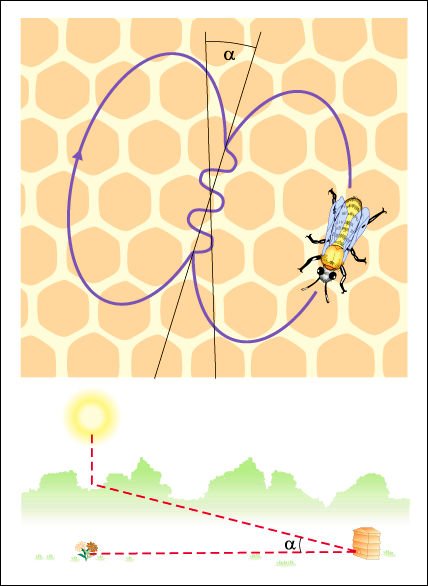
A medida que el Sol se mueve, el ángulo de la danza rota hacia el oeste.
Las danzas estereotipadas se
producen sobre panales ubicados en posición vertical. Se ha demostrado
que el ángulo que forma el eje de la danza con la vertical se correlaciona
con el ángulo formado entre la dirección del Sol y la de la fuente
de alimento.
En la danza del contoneo -o
danza en ocho-, la obrera recolectora que regresa de una fuente de alimento
realiza una caminata recta sobre el panal, mientras mueve su abdomen hacia los
lados. Luego de este recorrido, camina en círculo y luego retoma la caminata
recta y repite el comportamiento anterior. Al mismo tiempo, produce un sonido
característico, aunque no transmite vibraciones. Este comportamiento
se repite varias veces y la trayectoria de la abeja toma la forma de un ocho.
Se demostró que la duración del sonido en la parte recta de la
danza -como así también la longitud de la trayectoria recorrida
y su duración- están correlacionadas positivamente con la distancia
de la fuente que, dependiendo de qué especie se trate, es en general
mayor de 90 metros
La danza circular es ejecutada
cuando la recolectora retorna de fuentes de alimento cercanas a la colmena.
En este caso, la danzarina realiza intensos movimientos en círculo a
favor y en contra de las agujas del reloj y emite sonidos sin generar vibraciones
a nivel del sustrato. La danza circular se podría interpretar como una
danza en ocho sin contoneo, que indicaría que la fuente de alimento está
muy cerca
Sociedades de vertebrados
Con raras excepciones, las sociedades
de vertebrados no tienen el rígido sistema de castas característico
de los insectos eusociales. Los estudios han revelado que muchas sociedades
de vertebrados están, no obstante, altamente estructuradas, con los papeles
sociales y el acceso a los recursos determinados por interacciones específicas
que varían de especie a especie y de acuerdo con la edad y el sexo de
los individuos.
En muchas especies de aves y
mamíferos, las jerarquías de dominancia, mantenidas por patrones
de comportamiento específicos de cada especie, determinan la prioridad
del acceso a los recursos e influyen fuertemente en el éxito reproductivo
relativo.
Las jerarquías de dominancia
pueden tomar la forma de órdenes de picoteo, donde los animales de elevado
rango tienen prioridad para acceder al alimento, a otros recursos y reproducirse
más. La territorialidad es un sistema de dominancia social, por el cual
sólo los animales con territorio se reproducen y los animales con mejores
territorio se reproducen más. Los territorios pueden ser "reales"
-es decir, pueden ser áreas reales que contengan alimento y material
con que construir los nidos para mantener a una pareja y a sus crías-
o pueden ser simbólicos, como una arena o una glorieta.
Habitualmente, sólo el
macho dominante y la hembra dominante se aparean, y el resto de la manada coopera
para cuidar a las crías. El cuidado de las crías incluye la vigilancia
de la guarida y la provisión de alimento, que es tragado al matar una
presa y luego regurgitado para las crías.
Muchos vertebrados permanecen
cerca de sus lugares de nacimiento, ocupando un área natal que probablemente
sea igual a la que ocuparon sus padres. Frecuentemente, estas áreas natales
son defendidas por individuos o por grupos, contra otros individuos o grupos
de la misma especie o de especies estrechamente emparentadas, que utilizan los
mismos recursos. Las áreas defendidas de esta manera se conocen como
territorios, y el comportamiento de defensa de un área contra rivales
se conoce como territorialidad.
A raíz de la territorialidad,
aumenta la probabilidad de que una pareja que se aparea obtenga alimento y material
para la construcción del nido en el área y un lugar seguro para
desarrollar las actividades asociadas con la reproducción y el cuidado
de las crías. Los territorios están claramente definidos, reconocidos
y defendidos por el propietario del territorio.
Selección por parentesco
En 1962, V. C. Wynne-Edwards
propuso que los individuos que no podían reproducirse lo hacían
en beneficio de la sociedad a la cual pertenecían.Este
comportamiento era perpetuado por la mayor supervivencia de los grupos cuyos
miembros se comportaban con esta continencia altruista.
Esta explicación, llamada
selección grupal, proponía que los animales estaban genéticamente
programados para frenar su reproducción, para el bien de la sociedad
en conjunto. Este concepto fue rechazado porque no había una manera de
demostrar que la selección grupal pudiese ser mantenida contra la selección
natural individual para incrementar la reproducción. Sin embargo, esta
propuesta sirvió para galvanizar toda una serie de estudios extremadamente
provechosos, que han revolucionado la forma en que los biólogos modernos
encaran el comportamiento social.
El rechazo de la selección
del grupo llevó a otra propuesta, elaborada por W. D. Hamilton y basada
fundamentalmente en estudios de los insectos sociales.
En genética de poblaciones,
la medida de la aptitud no es el número de progenie que sobrevive, sino
el incremento o disminución de alelos particulares en el reservorio génico.
Una madre correría riesgos
y haría sacrificios por su prole de manera que ésta incremente
la representación de sus alelos en el reservorio génico. En la
medida en que el correr estos riesgos o el hacer estos sacrificios incremente
su contribución al reservorio génico, los alelos que dictan este
programa de correr riesgos se incrementarán en la generación siguiente,
y así sucesivamente. Pero, los hermanos que tienen el mismo padre comparten
unos con otros, en promedio, la mitad de sus alelos. Por lo tanto, cualquier
alelo que influyera favorablemente en el comportamiento altruista entre los
hermanos podría incrementar de manera similar su representación
en el reservorio génico.
La hipótesis de Hamilton
basada en este principio se llama selección por parentesco, que se define
como la reproducción diferencial de grupos diferentes de individuos emparentados
de una especie que se reproducen con tasas diferentes. El factor crítico
en la selección por parentesco es el efecto del individuo sobre el éxito
reproductor de sus parientes. Las pruebas de la hipótesis están
basadas en los grados de vinculación entre los individuos. Es posible
calcular, con relativa facilidad, el grado de parentesco entre dos individuos.
Por ejemplo, los hermanos, al igual que la mayoría de los otros hermanos
que tienen el mismo padre, poseen en promedio el mismo grado de parentesco unos
con otros que con sus padres y con su propia progenie.
Hamilton usó su hipótesis
para explicar la evolución de castas estériles entre las abejas.
Aunque esta explicación acerca de la selección por parentesco
entre las abejas sufrió ciertas críticas, sirvió para establecer
una nueva perspectiva evolutiva, la de la aptitud inclusiva. El criterio de
aptitud darwiniana se refiere al número relativo de descendientes del
individuo que sobreviven para reproducirse. El criterio de aptitud inclusiva
se refiere al número relativo de alelos de un individuo que pasan de
una generación a otra, ya sea como resultado de su propio éxito
reproductivo o del de los individuos emparentados. La hipótesis de Hamilton
es valiosa porque es verificable, es decir, permite hacer predicciones, probarlas
en un experimento y según los resultados, aceptaras o rechazarlas.
El gen egoísta
Algunos biólogos ven
a un organismo simplemente como el medio que un gen utiliza para producir más
genes. El argumento es simple: el organismo individual es transitorio. El genotipo
se fragmenta cada generación. Todo lo que puede sobrevivir de una generación
a otra es el gen. La manera de cómo sobrevive es en forma de réplicas.
Cuanto más réplicas, mayor será la probabilidad de que
sobreviva. El organismo es la máquina de supervivencia del gen y, por
lo tanto, programa a la máquina de tal modo que produzca copias de los
genes a la velocidad máxima, independientemente del costo personal para
el organismo.
Actualmente hay acuerdo general
en que el concepto de gen egoísta es una simplificación exagerada,
como opina incluso su padrino, Richard Dawkins. Los científicos opositores
a este concepto se basan en que, en la actualidad, se sabe que los genes no
actúan solos sino que están en permanente interacción y
que, además, presentan jerarquías de dominancia. Los organismos
no están determinados por sus genes, aunque sí están influidos
por ellos. Para comenzar a discutir estas ideas es necesario entender los factores
involucrados en el desarrollo de un organismo: el desarrollo no depende sólo
de los materiales que fueron heredados de los padres -como genes y materiales
contenidos en los gametos- sino de factores ambientales como temperatura, humedad,
nutrición, olores, sonidos y -en humanos- lo que se denomina educación.
Las variaciones entre individuos
de una misma especie son una consecuencia de la constante interacción
entre los genes y el ambiente en el que el organismo se desarrolla. Pero además,
Lewontin menciona un factor adicional: la variación al azar que se produce
durante el desarrollo y que provoca, por ejemplo, que organismos genéticamente
idénticos, criados en las mismas condiciones, presenten diferencias fenotípicas
marcadas.
Al dirigir su atención a la supervivencia de los genes antes que a la de los individuos, los biólogos que apoyaban esta postura comprendieron que existían oportunidades para conflictos serios de interés, no sólo entre individuos que se encuentran en competencia directa por un recurso limitado sino también entre los miembros del núcleo familiar. Entre los conflictos de interés librados con mayor ardor, están los que ocurren entre los miembros de una pareja reproductora.

Conflicto entre dos peces de franjas azules en el límite de sus territorios. Cada uno aferra al otro por los labios y el más débil es el primero que se aleja. Aunque vencido, el perdedor vuelve indemne a su propio territorio.
Los gametos femeninos son relativamente costosos, en términos metabólicos,
y si la hembra también debe llevar al embrión en su cuerpo y cuidar
de las crías después del nacimiento, su inversión en cada
esfuerzo reproductivo puede ser realmente muy grande, considerando su potencial
reproductor total. Los gametos masculinos, en contraste, habitualmente son "baratos":
una sola inseminación aporta gran cantidad de espermatozoides con relativamente
bajo costo. En la mayoría de las especies animales, toda la contribución
del macho a la generación siguiente consiste en sus genes. En estas situaciones,
interesa a los genes "egoístas" de la hembra encontrarse en
la "mejor compañía posible", promoviendo de este modo
su supervivencia en la generación siguiente. De aquí, puede surgir
una intensa competencia entre los machos en la que demostrarán a las
hembras que son los mejor dotados.
Altruismo recíproco
Los actos aparentemente altruistas
pueden tener sentido porque incrementan la probabilidad de supervivencia de
genes compartidos por el que realiza el acto y el beneficiario. Existe otro
modelo para un comportamiento aparentemente de autosacrificio, que se llamó
altruismo recíproco. De acuerdo con este modelo, un acto altruista es
llevado a cabo con la expectativa de que se retribuya el favor.
Para que el altruismo recíproco
sea una estrategia exitosa, resistente al fraude, es necesario que los individuos
sean capaces de reconocerse mutuamente y que un individuo coopere con otro individuo
o lo engañe, sobre la base de lo que hizo este último la ocasión
anterior en que ambos se encontraron.
La biología de la conducta
humana
Es tentador -en verdad, casi
irresistible- trazar paralelismos entre el comportamiente de los humanos y el
observado en otras especies. Hasta qué punto estos conceptos concernientes
a la evolución del comportamiento pueden extrapolarse a la especie humana,
es una cuestión que se debate actualmente.
Un grupo de biólogos
sostiene que la especie humana básicamente no es diferente de cualquier
otra especie y que si buscamos modificar la conducta humana para el bienestar
común, debiéramos comprender sus raíces.
Un grupo de opositores sostiene
que mientras los tempranos antecesores humanos pueden haber sido gobernados
por sus genes en el pasado, los humanos modernos son también producto
de su cultura y de su experiencia individual y, por consiguiente, aquellos análisis
ya no son válidos. Además, presentan otros argumentos, como los
que se basan en los factores que afectan el desarrollo de un individuo, ya mencionados
en oposición al concepto del gen egoísta.
Muchos científicos consideran
que hay que ser cauteloso cuando se considera que la biología determina
la conducta humana ya que estas ideas yacen en las raíces de todas las
nociones de superioridad racial. Así, han proporcionado la fundamentación
para la esclavitud, la explotación y el genocidio.
Más comúnmente,
la noción de que nuestra conducta está determinada, hasta cierto
grado, genéticamente, nos permite perdonarnos a nosotros mismos por la
violencia, la agresividad, la docilidad y la codicia, e inclusive, justificarlas.


