Capítulo 26.
La clasificación
de los organismos
La mayoría de las personas
tienen un conocimiento limitado del mundo natural y se relacionan principalmente
con los organismos que influencian sus propias vidas. Los biólogos se
enfrentan con la enorme tarea de clasificar, determinar e intercambiar información
acerca de la vasta diversidad de organismos con la que los seres humanos, recién
llegados en un sentido evolutivo, compartimos el planeta. Para esto, los biólogos
deben disponer de un sistema de clasificación que les permita nombrar
y agrupar a las especies descriptas de una manera lógica, objetiva, económica
y no redundante. La construcción de un sistema como éste no es
trivial si consideramos que, como mínimo, existe un número de
especies sin clasificar similar al número de especies ya descriptas -alrededor
de 1 millón y medio-. Por siglos, los naturalistas han intentado describir
y explicar la diversidad del mundo natural. A esta tarea se la ha denominado
sistemática .
Designadas con un nombre genérico
y un adjetivo modificador, las especies son las unidades básicas de clasificación
biológica. Aunque en latín especie simplemente significa "tipo"
y, por lo tanto, en el sentido más simple, las especies son tipos diferentes
de organismos, se utiliza el término especie en sentidos distintos.
El área el conocimiento
encargada de establecer las reglas de una clasificación es la taxonomía.
De este modo, la sistemática biológica utiliza la taxononomía
para establecer una clasificación.
La clasificación debe
representar en buena medida la filogenia de todos los seres vivos que han surgido
en este planeta. La sistemática evolutiva intenta no sólo hacer
buenas clasificaciones sino hacerlas de manera objetiva y sin arbitrariedades.
La filogenia de un grupo de especies cualesquiera puede representarse en forma
de árbol ramificado. Este tipo de diagrama representa una hipótesis
de las relaciones de ancestralidad y descendencia de las especies que contiene.
La teoría sistemática
se ha nutrido del aporte y discusión de taxónomos de diferentes
escuelas: la de los feneticistas, los cladistas y los evolucionistas. En este
sentido, las clasificaciones en clados , sólo interesadas en representar
las relaciones de ancestralidad y descendencia, son a las que adhieren la mayor
parte de los biólogos en la actualidad.
La sistemática molecular
ha ido en busca de grandes cantidades de similitudes homólogas con el
desarrollo de numerosas técnicas: la secuenciación de proteínas
, de ácidos nucleicos y otras técnicas moleculares. El descubrimiento
de moléculas y regiones de DNA que registran el cambio evolutivo a distintas
tasas ha permitido transformar la sistemática clásica en una sistemática
universal.
Con el desarrollo del microscopio
se descubrieron una gran cantidad de microorganismos y su clasificación
se hacía cada vez más necesaria. Hasta hace poco tiempo, el reino
se consideraba la categoría sistemática más inclusiva.
Sin embargo, la secuenciación de moléculas universales -presentes
en todos los organismos- llevaron a algunos científicos a la construcción
de un árbol filogenético único en el cual se diferencian
tres linajes evolutivos principales. Se propuso entonces la categoría
de dominio para cada uno de estos linajes, o grupos monofiléticos , y
los denominó Bacteria, Archaea y Eucarya.
La clasificación en reinos
y dominios se encuentra en movimiento cambiante permanente. La discusión
acerca de la validez de las clasificaciones nos hace reflexionar acerca de la
facilidad con la que solemos argumentar a favor de hipótesis cargadas
con valoraciones humanas, como el incremento de complejidad y el progreso evolutivo.
Las clasificaciones cladísticas, aunque puedan narrar historias evolutivas
incompletas en términos biológicos, son hipótesis objetivas
y comprobables en cualquier rango de la jerarquía biológica.
La necesidad de una clasificación
Hay aproximadamente un millón
y medio de especies descriptas y se cree que este número representa sólo
el 5% de las especies con las que actualmente compartimos el planeta. Durante
siglos, los naturalistas se han interesado en ordenar esta diversidad y, al
hacerlo, surgió un patrón jerárquico como norma de la clasificación
biológica.
Las especies se agrupan en géneros,
los géneros en familias, las familias en clases, las clases en órdenes,
los órdenes en phyla, los phyla en reinos y éstos en dominios.
La posibilidad de utilizar esta clasificación inclusiva de grupos dentro
de grupos es otra evidencia más a favor del proceso de evolución
de las especies.
¿Qué es una especie?
Una definición rigurosa
de especie (aunque no es la única) fue propuesta por Ernst Mayr, biólogo
evolutivo de la Universidad de Harvard, en 1940. Bajo el título de especie
biológica, Mayr describió a una especie como "un grupo de
poblaciones naturales cuyos individuos se cruzan entre sí de manera real
o potencial y que están reproductivamente aislados de otros grupos".
La expresión "real
o potencial" tiene en cuenta el hecho de que, aunque es improbable que
individuos de poblaciones geográficamente aisladas se crucen naturalmente,
el traslado de un grupo de organismos a alguna isla remota no los convierte
automáticamente en miembros de una especie distinta ya que éstos
potencialmente pueden cruzarse. La especiación requiere el establecimiento
de una o varias barreras que aseguren el aislamiento reproductivo. Los términos
"grupos" y "poblaciones" también son importantes
en esta definición. La posibilidad de que algunos individuos de especies
diferentes tengan una progenie ocasional no es relevante como proceso natural
si no conviven en el mismo habitat natural.
Si no existiesen barreras de
aislamiento reproductivo entre especies distintas, los organismos de una especie
podrían intercambiar genes con los miembros de otra especie y, en consecuencia,
no retendrían las características morfológicas, comportamentales
y genéticas que los identifican como tipos diferentes de organismos.
El término "especie"
tiene tres usos distintos. Hablamos de especie cuando nos referimos a la categoría
o rango taxonómico de especie. También hablamos de especie cuando
nos referimos a un taxón, es decir, a una clase lógica formada
por individuos que agrupamos en virtud de ciertos atributos comunes que un taxónomo
ha definido con anterioridad.
Finalmente, el término
especie también hace referencia a las unidades evolutivas que habitan
el mundo natural y a las que E. Mayr ha definido por medio del concepto biológico.
La especie como taxón y la especie biológica - o bioespecie- no
deberían representar entidades distintas. Las primeras representan hipótesis
que, por medio de definiciones cada vez más ajustadas, intentan acercarse
a las segundas. De este modo, una especie es una categoría, una hipótesis
perfectible y también un concepto biológico. Mientras que la categoría
y el taxón especie se encuentran definidos en el campo del conocimiento
humano, o ámbito gnoseológico, la especie biológica se
define en el mundo real, o ámbito óntico, y nos advierte que hay
algo allí afuera, una entidad natural, que merece ser llamada especie.
De acuerdo con el sistema binomial
de nomenclatura, ideado por el naturalista sueco Linné (Linneo) en el
siglo XVIII, el nombre científico de un organismo está formado
por dos partes: el nombre genérico y un epíteto específico
(un adjetivo o modificador). Por convención, los nombres del género
y de la especie se escriben en letra cursiva. El nombre del género siempre
antecede al epíteto -Drosophila melanogaster- y solamente puede utilizarse
sin él en los casos en los que nos referimos al conjunto total de especies
que constituyen ese género, como cuando mencionamos a Drosophila,
Paramecium o Viola.
Clasificación y jerarquía
La taxonomía permite
organizar la diversidad de cualquier conjunto de objetos, ya sean libros de
una biblioteca, víveres de una estantería o las especies de un
ecosistema. Cuando se aplican ciertas reglas de clasificación a los seres
vivos, se genera un sistema jerárquico, es decir, un sistema de grupos
dentro de grupos. La clasificación jerárquica no es una consecuencia
natural de la aplicación de reglas taxonómicas a cualquier sistema
de objetos. La naturaleza jerárquica de la clasificación biológica
surge como una consecuencia del proceso de evolución de las especies.
En la época de Linneo,
existían tres categorías básicas: la especie, el género,
y el reino. Los naturalistas reconocían 2 reinos biológicos: vegetal
y animal. Posteriormentre, el mismo Linneo y otros taxónomos fueron añadiendo
categorías intermedias entre género y reino. Los géneros
fueron agrupados en familias, las familias en órdenes, los órdenes
en clases y las clases en phyla o divisiones. Estas categorías pueden
a su vez subdividirse o agruparse en otras menos frecuentes como tribus, superfamilias
o subphyla. Muchos biólogos reconocen hoy una categoría por encima
del reino, el dominio. Para determinar que un individuo pertenece a una especie,
se requiere una gran cantidad de información. Una clasificación
jerárquica es una manera económica de manejar la información
biológica.
En el sistema jerárquico
de clasificación biológica, cada grupo o taxón tiene asociado
una categoría y un conjunto de atributos que determina la pertenencia
de ciertos organismos a ese grupo.
Las categorías y los
taxa de cualquier rango, no solamente el de especie, son sólo construcciones
mentales, sin embargo, esto no habilita a los taxónomos a formular cualquier
tipo de clasificación.
Los taxónomos han discutido durante mucho tiempo las virtudes y falencias de distintos métodos de clasificación. Finalmente, una idea se ha generalizado: si se pretende llegar a una clasificación objetiva, ésta debería ser única y, si es única, nada mejor que represente la historia evolutiva de los organismos que viven y han vivido en este planeta; una historia que, sin duda, será irrepetible.
La naturaleza jerárquica de la clasificación biológica consiste en la formación de grupos dentro de grupos.
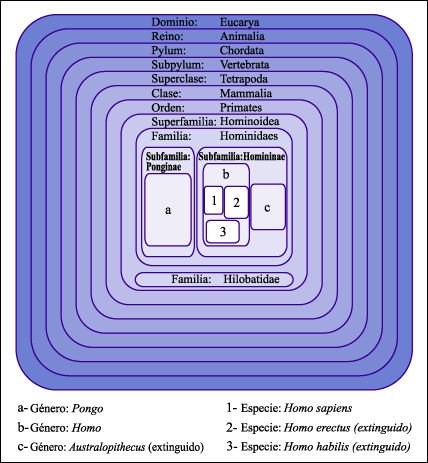
Los grupos, también llamados inclusivos, pueden representarse en un diagrama
de Venn. Este diagrama muestra la clasificación reciente del género
Homo. Note que el taxón de rango familiar Hominidae, que hasta hace poco
incluía solamente al género Homo, actualmente se propone como
un grupo monofilético que contiene a las especies de los géneros
Pongo (orangutanes), Gorilla (gorilas), Pan (chimpances) y Homo (humanos). La
subfamilia Homininae contiene, a la vez, a estos tres últimos taxa.
Sistemática y evolución
Las similitudes entre organismos
pueden constituir analogías u homologías, respectivamente, y su
distinción es la clave para la formación de grupos inclusivos.
Un ejemplo clásico de
homología lo constituye el miembro anterior de los tetrápodos.
El ala de un ave, la aleta de una ballena, la pata de un caballo y el brazo
de un hombre, a pesar de tener funciones distintas como volar, nadar, correr,
o agarrar, comparten un mismo patrón estructural: todas estos miembros
están formados por los mismos tipos de huesos (un húmero, un radio,
un cúbito, una serie de metacarpales y, en términos generales,
cinco dígitos).
Las estructuras que tienen un
origen común, pero no necesariamente conservan la misma función,
se denominan homólogas y constituyen una evidencia a favor de la hipótesis
de que estas seis especies derivan de un mismo ancestro común.
Los distintos huesos de las
extremidades anteriores de los animales de la figura se muestran en color para
indicar las similitudes fundamentales de estructura y organización.
Esta similitud apoya la hipótesis que propone que todos los tetrápodos compartimos un antecesor común. Contrariamente, la forma fusiforme de un pez y la de un delfín son similitudes análogas ya que, muy probablemente, la selección natural operando independientemente en dos linajes distintos haya beneficiado a los individuos que minimizaron la fricción y agilizaron su locomoción en el agua. Mientras que la homología nos permite distinguir relaciones de ancestralidad y descendencia, las analogías son un problema al momento de reconocer similitudes compartidas por una historia evolutiva en común. Si se pudiese agrupar a toda la diversidad de organismos vivientes y extinguidos por medio de similitudes homólogas, la clasificación representaría en buena medida la filogenia de todos los seres vivos que han surgido en este planeta.
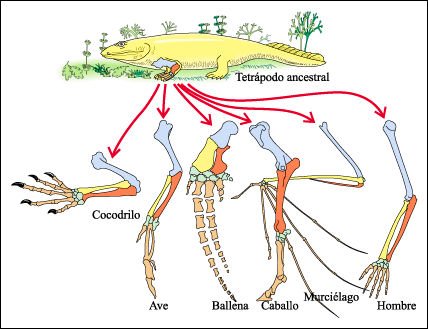
Huesos de extremidades anteriores que muetran las similitudes fundamentales de estructura y organización en diferentes animales.
La filogenia de un grupo de especies cualesquiera puede representarse en forma
de árbol ramificado. Este tipo de diagrama representa una hipótesis
de las relaciones de ancestralidad y descendencia de las especies que contiene.
Si se quiere clasificar a una nueva especie, el taxónomo debe previamente
construir un árbol filogenético, proponer una ubicación
coherente para la nueva especie y, posteriormente, derivar una clasificación
lógica. Para que la clasificación refleje con precisión
las relaciones de ancestralidad y descendencia, los taxa deben cumplir una única
condición, ser estrictamente monofiléticos. Esto significa que
todos los miembros de un taxón, cualquiera sea su categoría, deben
ser descendientes de una única especie, la especie ancestral más
próxima a todas las que contiene ese taxón.
Así, un taxón
genérico debe contener exclusivamente a las especies que han descendido
del ancestro común más cercano a todas las especies que contiene
ese género. Una familia debe contener sólo a los géneros
que han derivado de la especie ancestral más cercana de todas las especies
que contiene esa famillia. Los taxa generados de este modo no son sólo
construcciones mentales, sino que representan unidades históricas, ya
que son poseedores de un pasado único, exclusivo e irrepetible. A estos
agrupamientos se los denomina clados.
Cuando una clasificación se hace de modo tal que no respeta la formación de grupos monofiléticos -por considerar que otros criteros de clasificación son más adecuados para sus propósitos sistemáticos- los agrupamientos taxonómicos que surgen de esa clasificación no corresponden a grupos históricos. En este caso, los taxa pueden ser parafiléticos o polifiléticos. Las características que surgen como adaptaciones convergentes a un mismo modo de vida generan problemas en el momento de formar grupos monofiléticos. La formación de taxa por convergencias evolutivas suele formar grupos polifiléticos. Por otra parte, la divergencia evolutiva extrema de un grupo de especies suele llevar a la formación de grupos parafiléticos.
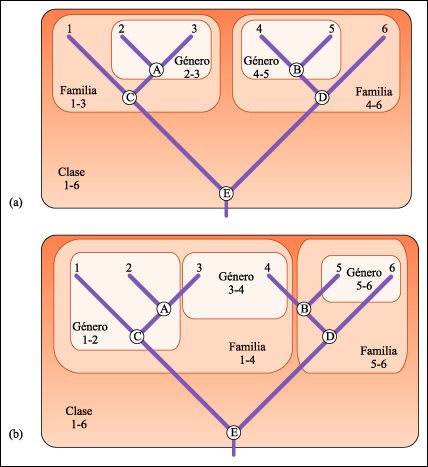
Distintos agrupamientos taxonómicos.
a) La formación de grupos monofiléticos se realiza luego de la
reconstrucción de las relaciones de ancestralidad y descendencia de un
grupo de especies. Los nodos (A-E) no representan a especies vivas o extinguidas,
sino al conjunto de caracteres que comparten las especies que descienden de
ese nodo. El nodo A contiene las características que comparten las especies
2 y 3, por ello, puede reconocerse un grupo monofilético denominado género
2-3. El nodo C contiene los caracteres compartidos por dos géneros distintos,
el género monoespecífico 1 y su grupo hermano, el género
2-3. Por ello, el taxón monofilético que contiene a todos los
descendientes del nodo C merece un rango más inclusivo, el de familia
1-3. Con la misma lógica, el nodo E contiene la serie de caracteres que
permiten reconocer a las seis especies como pertenecientes a un mismo grupo
histórico, la clase 1-6. b) Cuando los taxa se forman en ausencia de
esta lógica, pueden ser parafiléticos, como en el caso de los
géneros 1-2, 5-6 y la familia 5-6; o polifiléticos, como en el
caso del género 3-4 y la familia 1-4. Observe que los grupos parafiléticos
contienen al ancestro de todas sus especies descendientes pero excluyen al menos
a uno de sus descendientes. Los polifiléticos, sin embargo, no contienen
al ancestro común de ninguna de sus especies.
Escuelas sistemáticas
Existen diferentes escuelas
que han realizado aportes a la teoría sistemática: la escuela
feneticista, la cladista y la evolucionista.
La escuela feneticista argumenta
que una clasificación es tanto más informativa cuanto mejor refleja
la similitud global de un grupo de especies. Los feneticistas no llaman a sus
árboles filogenias, sino fenogramas, ya que reconocen que el parecido
fenotípico de un grupo de especies puede no representar una ancestralidad
común. Los feneticistas incluyen en sus clasificaciones a grupos monofiléticos,
parafiléticos y polifiléticos. A diferencia del cladismo y el
evolucionismo, la taxonomía numérica desarrollada por los feneticistas
sirve para agrupar, por parecido global, cualquier sistema de objetos, ya sean
biológicos o no biológicos. Las técnicas fenéticas
han servido a los genetistas, antropólogos y lingüistas para construir
una historia hipotética acerca de la diferenciación de las poblaciones
humanas .
El cladismo, o sistemática
filogenética, sostiene que las clasificaciones biológicas deben
representar un único proceso, la formación de linajes independientes
a partir de un ancestro común. Para los cladistas, los taxa deben consistir
sólo de grupos monofiléticos. Los taxa parafiléticos y
los polifiléticos no representan unidades históricas, por lo que
no pueden considerarse taxa válidos. Los cladistas construyen las clasificaciones
a partir de la deducción previa de un cladograma. Este tipo de clasificación
es un reflejo exacto del orden de ramificación o formación de
especies nuevas en el curso de la evolución. A diferencia de los feneticistas,
los cladistas no consideran relevante para una clasificación biológica
el cambio acumulado en un mismo linaje o rama del árbol filogenético.
Los cladistas se basan en el
reconocimiento de sinapomorfías. Si los taxa se construyeran por simplesiomorfías
sería imposible generar grupos inclusivos.
La escuela evolucionista ha
sostenido durante años que una clasificación debe considerar tanto
las relaciones de parentesco como la similitud fenotípica global, evitando
los agrupamientos polifiléticos. De este modo, las clasificaciones evolucionistas
pueden reconocer tanto a grupos monofiléticos como parafiléticos.
La divergencia morfológica extrema de un linaje como consecuencia de
la conquista de un nuevo nicho ecológico debe, según los evolucionistas,
estar reflejada en la clasificación.
Las propuestas clasificatorias
de las distintas escuelas no son adecuadas para todos los propósitos
sistemáticos. Aunque la escuela cladista presenta una consistencia lógica
más robusta y menos subjetiva que las otras, no hay que olvidar que sus
clasificaciones son hipótesis históricas realizadas sobre un conocimiento
incompleto del mundo natural y que, como en cualquier disciplina científica,
esas hipótesis son perfectibles.
Sistemática molecular
La reconstrucción filogenética
y la clasificación sistemática dependieron, durante años,
de la experiencia, la sabiduría y el buen criterio de especialistas dedicados
al estudio de grupos específicos. A partir de la década de 1960,
distintas técnicas moleculares sirvieron para la reconstrucción
filogenética y la clasificación sistemática.
Una de las primeras proteínas
analizadas en los estudios de sistemática molecular fue el citocromo
c, una proteína de la cadena de transporte de electrones. Esta molécula,
secuenciada en una gran variedad de organismos, permitió determinar el
número de aminoácidos en que diferían cualquier par de
especies.
Cuanto mayor es el número de aminoácidos distintos, mayor debe haber sido el tiempo de evolución a partir de su ancestro común; inversamente, cuanto menor es el número de diferencias, más cercana debe haber sido la divergencia.
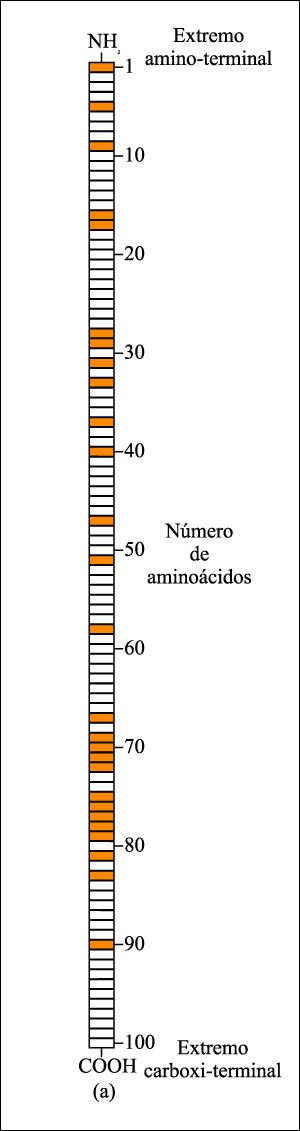
En las moléculas de citocromo c de más de 60 especies que han sido estudiadas, 27 de los aminoácidos son idénticos (color).
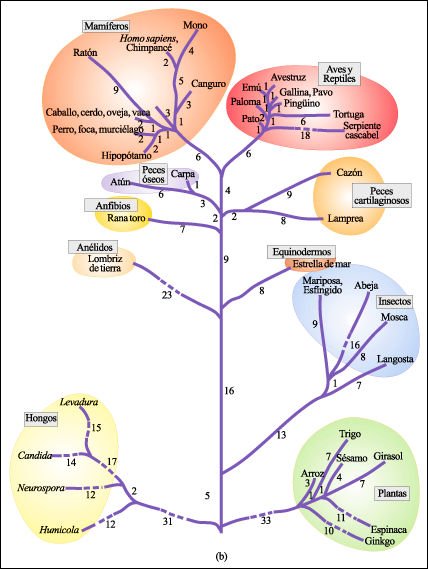
Las principales ramas del árbol filogenético basadas en las comparaciones de las secuencias de aminoácidos de las moléculas de citocromo c.
Los números indican la cantidad de aminoácidos por los cuales
cada citocromo c difiere del citocromo c correspondiente al punto de bifurcación
más cercano. Las líneas de guiones indican que se ha acortado
una rama y que, por lo tanto, no está representada a escala.
A medida que se acumularon datos
sobre la variación de distintas proteínas en diferentes organismos,
una corriente de biólogos evolutivos -los neutralistas- sugirió
que los cambios de aminoácidos en una proteína ocurren mayoritariamente
al azar y a una tasa constante. Los neutralistas sostienen que las distintas
formas de una misma proteína (dada por su secuencia de aminoácidos)
no han evolucionado por selección natural. Las variantes alélicas
de una misma proteína responden a la acumulación de cambios en
la secuencia de aminoácidos que resultan neutros, es decir, que no otorgan
ventajas ni desventajas selectivas. La variación neutra de una misma
proteína representa una manera útil de registrar el paso del tiempo.
Si las proteínas acumulan cambios a una tasa constante, los cambios pueden
considerarse como el tic-tac de un reloj. Este "reloj molecular" funciona
más rápido en las regiones de la proteína menos comprometidas
con su función.
Una vez calculada la tasa de
evolución de una proteína para un conjunto de especies de las
cuales se tiene registro de su tiempo de divergencia, se puede calcular el momento
en que han divergido dos especies cualesquiera. Esto se hace a partir de la
construcción de una curva de calibración. La calibración
del reloj molecular para una proteína específica se hace por medio
del uso de fósiles. Esto sirve para conocer el tiempo de divergencia
entre especies que no han dejado registro en las rocas.
Con el advenimiento de la técnica
de secuenciación de ácidos nucleicos, el uso de la secuenciación
de proteínas para estimar relaciones evolutivas fue abandonado. Esto
se debió, entre otras cosas, a que la secuenciación de ácidos
nucleicos es técnicamente mucho más fácil -ya que trata
solamente con cuatro nucleótidos diferentes comparado con los veinte
aminoácidos-. Además, una vez conocida la secuencia del DNA o
RNA, fácilmente puede deducirse la secuencia de proteínas.
El estudio de distintos tipos
de secuencias de DNA y RNA ha permitido encontrar relojes que funcionan a tasas
muy altas, muy bajas, e intermedias. Esta diversidad de tasas permite conocer
de manera aceptable las relaciones filogenéticas de distintos conjuntos
de organismos, relaciones entre padres e hijos, entre organismos de una misma
población, entre poblaciones y subespecies, entre especies aisladas reproductivamente
y entre taxa de rango supraespecífico, desde géneros distintos
hasta especies de dominios diferentes.
La clasificación de los
reinos y los dominios
Luego de la publicación
del Sistema Natural de Linneo en 1758, y durante muchos años, se reconocían
sólo dos ramas en la sistemática: la zoología y la botánica.
El evolucionista alemán Ernst Haeckel propuso, a finales del siglo pasado,
la construcción de un tercer reino, el de los Protistas, constituido
por microorganismos. Haeckel reconoció que algunos de estos microorganismos
carecían de núcleo celular y los denominó Monera. Posteriormente,
las bacterias fueron reconocidas, en 1956, por Herbert Copeland como reino Monera,
independiente de los Protistas. Los hongos, fueron los últimos organismos
que merecieron la creación de un reino y su fundador, R. Whittaker propuso,
en 1959, una clasificación general de los seres vivos que contenía
cinco reinos: Monera (bacterias), Protista (protozoos), Fungi (hongos), Animalia
(animales) y Plantae (plantas). Posteriormente, en 1978, Whittaker y Margulis,
propusieron una modificación, conservando el número de reinos
e incluyendo dentro del antiguo grupo Protistas a las algas. Este nuevo reino
fue denominado Protoctista; sin embargo, gran parte de la literatura científica
aún utiliza la denominación Protista. Así, esta nueva clasificación
de cinco reinos consiste en Procariota (bacterias), Protoctista o Protista (algas,
protozoos, mohos del limo, y otros organismos acuáticos y parásitos
menos conocidos), Fungi (líquenes y hongos), Animalia (animales vertebrados
e invertebrados) y Plantae (musgos, helechos, coníferas y plantas con
flor).
Hasta 1977, el reino se consideraba la categoría sistemática más inclusiva. Sin embargo, la secuenciación de moléculas universales que cambian a tasas extremadamente bajas (como en el caso del rRNA) llevaron a Carl Woese y sus colaboradores a la construcción de un árbol filogenético único en el cual se diferencian tres linajes evolutivos principales.
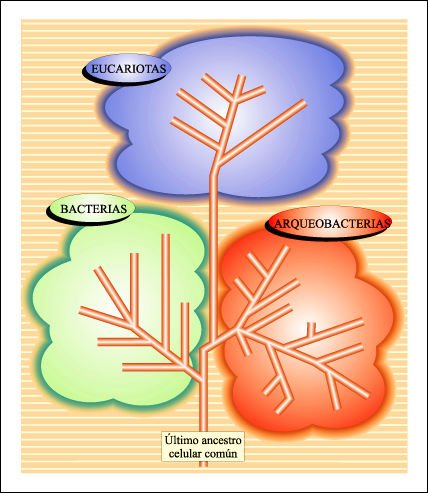
La estructura filogenética más profunda de la diversidad biológica obtenida por Carl Woese a partir de la secuenciación de rRNA.
En la clasificación de la figura anterior, claramente se distinguen tres
grupos monofiléticos distintos que corresponden a los dominios Bacteria,
Archaea y Eucarya.Woese propuso entonces la categoría de dominio para
cada uno de estos linajes, o grupos monofiléticos, y los denominó
Bacteria, Archaea y Eucarya. El cambio propuesto por Woese resalta las diferencias,
hasta ahora ocultas, entre organismos procariotas. De este modo, Monera es un
grupo parafilético que debería descartarse de la clasificación
biológica. En el sistema de Woese, Archaea y Bacteria son dominios distintos
de organismos procariotas y el primero contiene al menos dos reinos nuevos:
Crenarchaeota y Euryarchaeota. El dominio Eucarya agrupa, según esta
clasificación, a los restantes reinos de organismos eucariotas.
La clasificación de Woese,
como cualquier clasificación cladística, se basa en el orden de
ramificación de los linajes durante el curso evolutivo. Sin embargo,
no todos los taxónomos acuerdan con este principio clasificatorio y las
disidencias se acentúan cuando se trata de los taxa más inclusivos
de la clasificación biológica. La propuesta alternativa de Margulis,
centrada en los recurrentes procesos de simbiosis, como la de Cavallier-Smith
en la que propone la categoría de imperio en lugar de dominio, representan
las principales propuestas evolucionistas alternativas a la cladística
de Woese.




