Capítulo 28.
Los
protistas
El reino Protista comprende una enorme variedad de organismos eucarióticos
, principalmente unicelulares, y además, algunas formas multicelulares.
Un factor principal en la evolución de los eucariotas puede haber sido
el establecimiento de relaciones simbióticas con células procarióticas
que, al ser incorporadas, finalmente se especializaron como mitocondrias y cloroplastos.
En el pasado, era común considerar a los protistas fotosintéticos
-las algas- como "plantas inferiores", y a los heterótrofos
unicelulares -los protozoos- como "animales inferiores". Análogamente,
los mohos mucilaginosos y los mohos acuáticos fueron considerados "hongos
inferiores". Sin embargo, en la actualidad resulta cada vez más
evidente que, exceptuando las algas verdes de las cuales se originaron las plantas,
los protistas actuales representan linajes que no están íntimamente
relacionados con aquellos que originaron a los miembros de los tres reinos multicelulares.
Así, los términos "algas" y "protozoos" fueron
abandonados como términos formales en la clasificación moderna;
sin embargo, persisten como términos informales.
Los protistas representan varios
linajes filogenéticos bastante distintos. Informalmente pueden ser agrupados
en autótrofos fotosintéticos (algas), unicelulares autótrofos
y/o heterótrofos (euglenidos y dinoflagelados), heterótrofos multinucleados
y multicelulares (mohos mucilaginosos y acuáticos) y heterótrofos
unicelulares (protozoos).
Los protistas autótrofos
fotosintéticos son sumamente importantes en la producción energética
global. Entre las algas, las diatomeas y las crisofitas son componentes importantes
del agua dulce y del fitoplancton marino.
Dentro de los protistas hay
grupos unicelulares flagelados que pueden incluir organismos fotosintéticos,
otros heterotróficos y algunos pueden incluir las dos formas de vida.
Entre ellos, los dinoflagelados se caracterizan por poseer dos flagelos que
baten en planos diferentes, haciendo que el organismo gire; los dinoflagelados
habitualmente tienen paredes de celulosa rígidas, de conformación
extravagante. Muchos de ellos son marinos, algunos son bioluminiscentes y otros
producen la conocida marea roja. Los euglenoideos -o euglenofitos- son un grupo
extremadamente numeroso que se encuentra principalmente en agua dulce pero también
los hay marinos, de aguas salobres y parásitos.
Los mohos mucilaginosos son
organismos heterótrofos y ameboides. Estos mohos se reproducen por la
formación de esporas . Hay dos grupos principales, los mixomicetes -o
mohos mucilaginosos plasmodiales, que son cenocíticos durante las etapas
no reproductivas- y los acrasiomicetes -o mohos mucilaginosos celulares, en
los cuales las células ameboides agrupadas retienen su identidad individual-.
Los mohos acuáticos -u oomicetes- son heterótrofos cenocíticos
que superficialmente se asemejan a hongos. Se reproducen tanto asexual como
sexualmente, sólo las esporas son flageladas y todos presentan oogamia.
Se piensa que los protistas
heterótrofos unicelulares -o protozoos- han evolucionado de antecesores
flagelados no fotosintéticos. Entre los protozoos se encuentran algunas
de las células más grandes conocidas y también las más
complejas. Tres grupos, los mastigóforos -o flagelados-, los sarcodinos
-o ameboides- y los cilióforos -o ciliados- incluyen tanto especies de
vida libre como parásitas y sus miembros pueden identificarse sobre la
base de sus estructuras locomotoras. Los opalínidos y los esporozoos,
en cambio, contienen sólo formas parásitas.
Los protistas exhiben una variedad
de respuestas por las que se desplazan frente a distintos estímulos ambientales.
Clasificación de los
protistas
Desde el siglo XIX, la palabra
protista remite a organismos unicelulares. Sobre esta base, diferentes autores
consideraron absurdo designar a las grandes algas marinas como protistas, ya
que éstas no son unicelulares ni tienen un tamaño reducido y prefirieron
denominar a este reino Protoctista. Si bien esta denominación resulta
más correcta, ya que no implica una limitación en el número
de células ni en el tamaño, lo cierto es que, en la práctica,
es muy poco utilizada.
Así como la denominación
de este grupo es conflictiva, no lo es menos su clasificación. En los
últimos tiempos, la clasificación de los protistas -así
como la de los procariotas - ha sufrido una considerable reestructuración
a medida que la microscopía electrónica y las modernas técnicas
moleculares y bioquímicas fueron aportando una enorme cantidad de nueva
información acerca de ellos. Esto provocó una revolución
en las antiguas clasificaciones, comparable a la ocurrida en la biodiversidad
animal y vegetal durante los siglos XVIII y XIX.
Cuestiones tales como el número de divisiones o phyla que constituyen el reino, las relaciones entre las varias divisiones y los phyla, la ubicación correcta de determinadas especies y los criterios que deben usarse para decidir al respecto son aún tópicos de discusión y de encendida controversia. En la actualidad, no existe una clasificación completa del reino protista que sea ampliamente aceptada por los biólogos que estudian estos organismos.
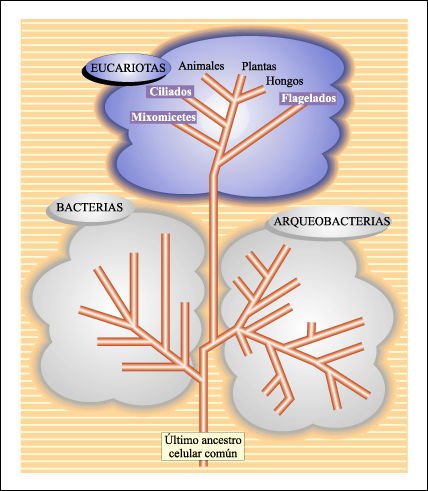
Esquema global de las relaciones evolutivas de los seres vivos.
Cabe recordar que los árboles como el de la figura anterior son hipótesis
y que, en la actualidad, se construyen a través de técnicas que
comparan secuencias de subunidades de rRNA. En este árbol se muestran,
en forma simplificada, sólo algunos representantes del reino Protista
debido a que no hay -al menos hasta el momento- un único árbol
que refleje las relaciones entre todos los miembros que conforman este grupo.
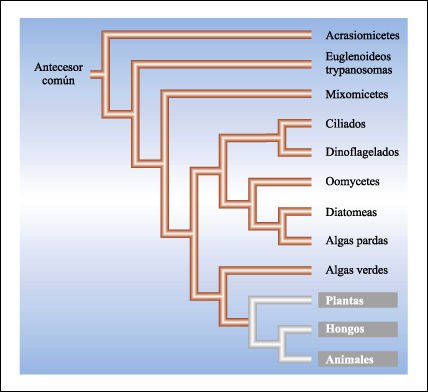
Posibles relaciones filogenéticas simplificadas entre los principales grupos de protistas.
Autótrofos fotosintéticos
A los autótrofos fotosintéticos,
con fines prácticos, se los denominan algas. Las algas, en general, tienen
una estructura relativamente simple; pueden ser unicelulares, un filamento de
células, una placa de células o un cuerpo sólido que puede
comenzar a aproximarse a la complejidad del cuerpo de una planta.
Las protistas unicelulares se
encuentran habitualmente flotando cerca de la superficie de océanos y
aguas continentales, donde abunda la luz.
La mayoría de las algas
multicelulares presentan adaptaciones que les permiten vivir en aguas poco profundas
a lo largo de las costas, donde se acumulan habitualmente muchos nutrientes
pero, las condiciones de vida son difíciles. Bajo estas presiones, distintos
grupos explotaron determinadas áreas a lo largo de las líneas
de marea, dando como resultado una zonación de formas de vidas diferentes,
características en muchas áreas costeras.
Las algas varían mucho
en sus características bioquímicas, especialmente en los pigmentos,
la naturaleza de las reservas alimenticias, los componentes de la pared celular
y en el número y posición de sus flagelos (cuando los tienen).
Típicamente, las paredes celulares de las algas tienen una matriz de
celulosa, pero algunas tienen otros polisacáridos que les dan una consistencia
mucilaginosa.
En un principio, los nombres
de algunas divisiones de algas derivaban de los colores de los pigmentos dominantes,
que enmascaran al verde brillante de las clorofilas. Al aumentar el número
de las divisiones, otros caracteres fueron usados para darles su nombre. La
gran variedad de pigmentos que se encuentran en los cloroplastos de las distintas
divisiones de algas sugiere que antes del desarrollo de las células eucarióticas
existían diferentes tipos de procariotas productores de oxígeno.
Luego, las distintas divisiones de algas podrían haber evolucionado como
resultado de relaciones simbióticas con los diferentes procariotas fotosintéticos,
los que posteriormente habrían originado a los actuales cloroplastos.
Entre las algas se encuentra también una gran diversidad de productos
de almacenamiento. La mayoría tiene reservas en forma de carbohidratos
y muchas contienen lípidos.
Los ciclos de vida de las algas
son extremadamente variados pero todas, con excepción de las algas rojas,
presentan células móviles flageladas, al menos en una etapa de
su ciclo de vida.
Las algas no constituyen un grupo natural dentro de los protistas sino que son un grupo polifilético.
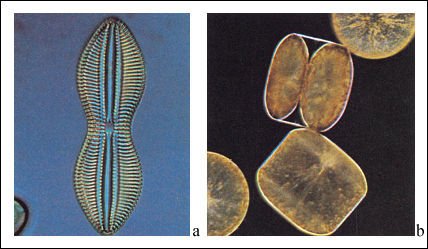
Diatomeas.
a) Visión lateral que muestra la valva pennada característica,
con dibujos intrincados. b) Vista desde arriba y de costado de una valva céntrica.
Nótese que una célula se está dividiendo. Cada célula
nueva recibirá una de las valvas y producirá la otra.
Las algas verdes interesan particularmente a los estudiosos de la evolución no sólo por sus relaciones con las plantas sino también por la amplia gama de complejidad que exhiben, ya que es un grupo que contiene organismos unicelulares y multicelulares. Una forma intermedia entre las formas unicelulares y las multicelulares está constituida por células individuales que se asocian en colonias. Las colonias difieren de los organismos multicelulares auténticos en que las células individuales preservan un alto grado de independencia. Frecuentemente están conectadas por cordones citoplasmáticos que unifican la colonia en grado tal que puede ser considerada como un sólo organismo. El ejemplo clásico de complejidad creciente entre los organismos que forman colonias se puede ver dentro del grupo de las volvocales. Dentro de ellas, la formación de colonias móviles está basada en la cohesión de células análogas a Chlamydomonas.
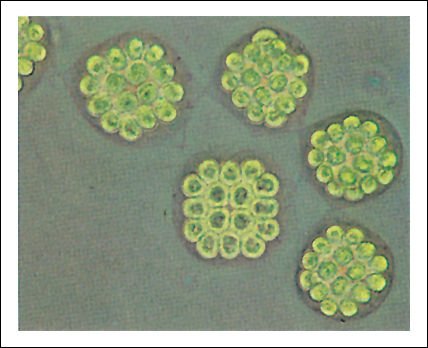
Volvocales: Gonium. Cada colonia está compuesta por una placa de células semejantes a Chlamydomonas, relacionadas por una matriz gelatinosa.
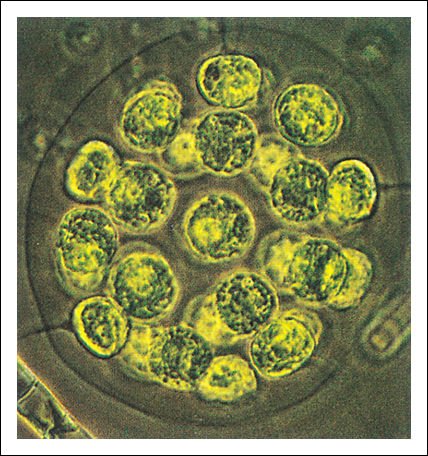
Pandorina, las células individuales forman una colonia en forma de huevo.
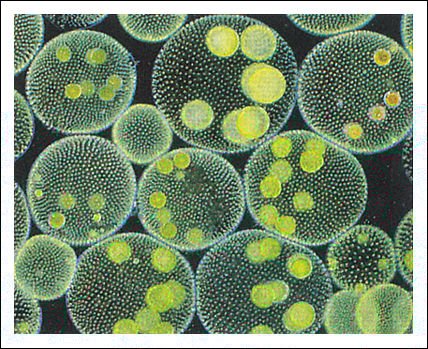
Volvox, cada colonia está constituida por centenares o miles (dependiendo de la especie) de células biflageladas, de color verde brillante, unidas entre sí por delgadas hebras de citoplasma.
Cada colonia de la figura anterior forma una esfera hueca que gira en el agua
como resultado del batir coordinado de los flagelos. Las colonias hijas se forman
dentro de la colonia madre; varias de ellas son visibles en esta fotomicrografía.
Si se comparan los géneros Chlamydomonas, Gonium, Pandorina y Volvox,
se puede observar una progresión continua en tamaño y complejidad,
y también una tendencia hacia la especialización de las funciones.
Sin embargo, este grupo particular representa un "callejón sin salida"
evolutivo, puesto que no ha originado a un grupo más complejo de organismos.Otro
tipo de forma de vida intermedia resulta luego de repetidas divisiones nucleares
que no son acompañadas de una división correspondiente del citoplasma
y la formación de paredes celulares.
Un organismo entero, como Valonia, puede parecer unicelular pero, de hecho, consiste en muchos núcleos dentro de un citoplasma común. Este tipo de organización multinucleada no es genuinamente ni unicelular ni multicelular, sino que se conoce como cenocítica.

Las clorofitas incluyen una variedad de organismos multinucleados y multicelulares además de las formas de las colonias.
a) Valonia , un alga verde cenocítica, aproximadamente del tamaño
de un huevo de gallina, contiene muchos núcleos, pero no tiene particiones
que los separen. Es común en las aguas tropicales. b) Spirogyra es un
alga de agua dulce en la cual las células se alargan y luego se dividen
por paredes transversales, de modo que pueden mantenerse juntas en filamentos
largos y finos. Los cloroplastos forman hélices que parecen bandas de
cinta verde dentro de cada célula. c) Ulva o lechuga de mar, es un alga
marina en la cual las células se dividen tanto longitudinal como lateralmente,
con una sola división en el tercer plano, produciendo un talo ancho.
Entre las algas verdes se observa una variedad de diferentes ciclos de vida. Chlamydomonas es unicelular haploide y se reproduce asexualmente.
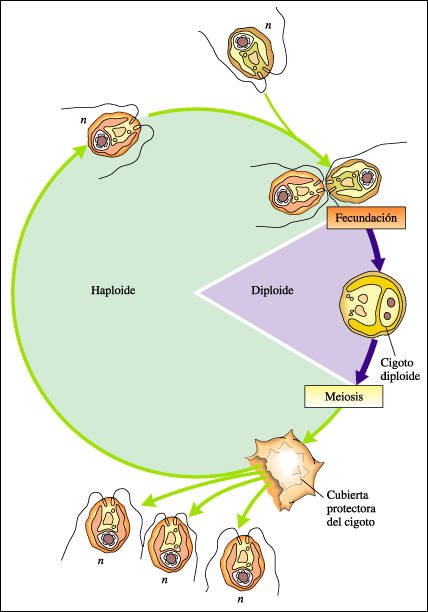
El ciclo vital de Chlamydomonas.
El organismo es haploide durante
la mayor parte de su vida. La fecundación se produce por la unión
de dos células fecundantes de cepas diferentes y da origen a un cigoto
diploide. El cigoto produce una cubierta gruesa que le permite permanecer latente
durante condiciones rigurosas. Después de este período de latencia,
el cigoto se divide por meiosis, formando cuatro células haploides. Cada
célula haploide puede reproducirse asexualmente (por mitosis) para formar
más células haploides o, en condiciones ambientales adversas,
las células haploides de una línea fecundante particular pueden
fusionarse con células de un tipo opuesto, iniciándose así
otro ciclo sexual.
Un ciclo de vida más complejo, caracterizado por la alternancia de generaciones, se encuentra en algunas algas verdes multicelulares, así como en todas las plantas.
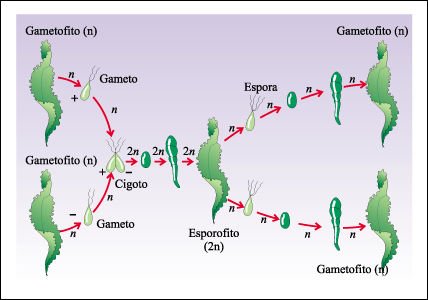
Ciclo de vida de la lechuga de mar.
La lechuga de mar, Ulva, presenta alternancia de generaciones en su ciclo de
vida, donde en una generación produce esporas y en la otra gametos. El
gametofito haploide (n) produce isogametos haploides que se fusionan y forma
el cigoto diploide (2n). El esporofito, cuerpo multinucleado formado por células
diploides, se desarrolla a partir del cigoto. Este produce esporas haploides
por meiosis, las cuales producen gametofitos haploides, y el ciclo comienza
nuevamente.
Protistas que presentan especies
autótrofas y heterótrofas
Durante los dos últimos
siglos se presentaron dificultades a la hora de clasificar organismos con características
intermedias entre plantas y animales, tales como movilidad, capacidad de realizar
fotosíntesis y heterotrofia. Organismos como los coanoflagelados, dinoflagelados
y euglenoides, indistintamente, fueron tratados como algas o como protozoos
o, lo que es peor aun, como ambos a la vez. Esto ha conducido a grandes problemas
de nomenclatura, que persisten hasta la actualidad.
Los dinoflagelados presentan una gran diversidad morfológica y funcional: en su mayoría son unicelulares, pero algunos forman colonias. Pueden ser fotosintéticos o heterótrofos (fagotrofos o parásitos), de vida libre o sésil y son componentes importantes del fitoplancton, tanto de aguas continentales como marinas. Muchos dinoflagelados presentan, por debajo de la membrana plasmática, vesículas que contienen placas de celulosa que le dan un aspecto rígido. El número, la forma y las ornamentaciones de las placas se utilizan para la determinación de las especies. También hay formas "desnudas", que incluyen algunas especies parásitas.
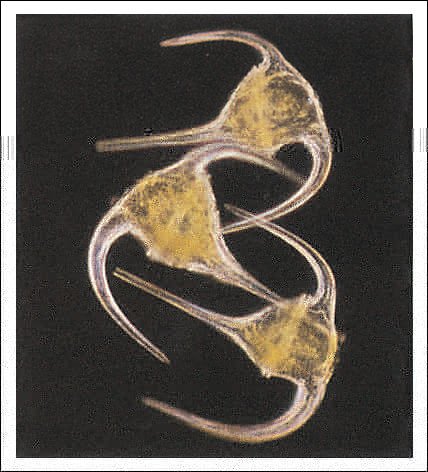
Dinoflagelados. Ceratium tripos, un dinoflagelado con placas.
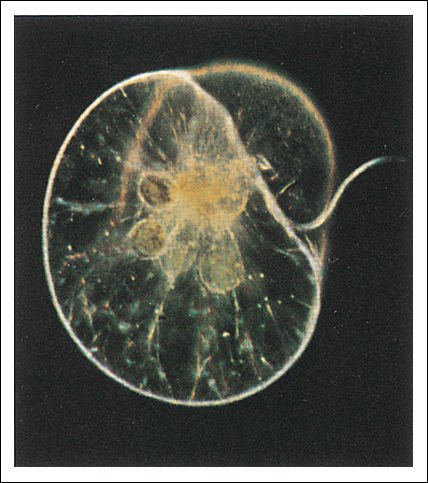
Noctiluca scintillans, un dinoflagelado marino bioluminiscente. Dentro de la célula pueden verse las diatomeas pardo-amarillas que han sido ingeridas.
Los euglenoides son un importante grupo de organismos unicelulares que pueden
vivir en agua dulce, salobre, marina, o en sedimentos húmedos; pueden
ser fotosintetizadores, heterótrofos o parásitos; de vida libre
o sésil. Reciben su nombre del género Euglena, el más conocido
del grupo.

Euglena es uno de los organismos unicelulares más versátiles. Contiene numerosos cloroplastos, realiza fotosíntesis, pero también puede absorber nutrientes orgánicos del medio circundante y algunas especies pueden vivir sin luz. Euglena nada por movimiento de su flagelo locomotor.
Heterótrofos multinucleados y multicelulares
Los mohos mucilaginosos son
un grupo de organismos curiosos, habitualmente clasificados con los protistas
a raíz de su similitud con las amebas. Los dos grupos principales que
se conocen son los plasmodiales o mixomicetes y los celulares o acraciomicetes.
La mayoría de estos mohos viven en lugares húmedos, fríos
y sombreados de los bosques; sin embargo, una de las especies plasmodiales comunes
(Physarum cinereum) se encuentra a veces deslizándose sobre céspedes
urbanos. Los mixomicetes presentan una variedad de colores.
Durante los estadios no reproductivos, los mixomicetes son masas delgadas y móviles de protoplasma, que avanzan lentamente a modo de amebas.
A medida que estos plasmodios
se desplazan, fagocitan bacterias, levaduras, esporas de hongos y pequeñas
partículas de materia vegetal y animal en descomposición que luego
digieren. Pueden crecer hasta pesar 50 gramos o más y, dado que se extienden
en forma de una película delgada, pueden llegar a cubrir un área
de más de un metro de diámetro.
El plasmodio es cenocítico;
a medida que crece, los núcleos se dividen repetidamente y, en las etapas
tempranas, sincrónicamente. Este crecimiento continúa en tanto
haya una reserva de alimentos y humedad adecuada. Cuando cualquiera de éstos
escasea, el plasmodio se divide en muchos montículos de protoplasma,
cada uno de los cuales produce un esporangio maduro -estructura en la cual se
desarrollan las esporas-, en el ápice de un pedicelo.
En el esporangio ocurre la meiosis y los núcleos haploides individuales que se forman son rodeados por paredes celulares. Esto da lugar a esporas que germinan cuando las condiciones son favorables; cada espora produce entre una y cuatro células haploides flageladas, según la especie. Algunas de estas células se fusionan y forman un cigoto a partir del cual se desarrolla un nuevo plasmodio.

Moho mucilaginoso.
En la figura anterior se puede observar: a) Un plasmodio de un moho mucilaginoso
en el que se ven múltiples núcleos. Este plasmodio puede pasar
a través de un trozo de seda o de un papel de filtro y aparecer del otro
lado, aparentemente sin cambio. b) esporangios de un moho mucilaginoso plasmodial
sobre un tronco en descomposición.
Los mohos acuáticos son
organismos cenocíticos, muchos de los cuales se asemejan a los hongos
en su estructura básica. Hasta hace muy poco se los solía clasificar
junto con ellos, pero sus características bioquímicas y la presencia
de células reproductoras flageladas (que no existen en ninguno de los
hongos) indica que representan un linaje distinto. El principal grupo son los
oomicetes. Algunos autores incluyen a los oomicetes dentro de un nuevo reino
denominado Stramenopila.
La mayoría de los mohos
acuáticos consisten en filamentos cenocíticos conocidos como hifas.
Como en los hongos, porciones de las hifas forman estructuras especializadas,
los gametangios, en donde se producen las gametas. Las paredes celulares de
los oomicetes, como las de muchas algas -pero a diferencia de las de los hongos-
contienen celulosa. Todos los oomicetes muestran oogamia. Sin embargo, los gametos
de los oomicetes no tienen flagelos y, por lo tanto, no son móviles.
En la reproducción sexual, el núcleo del espermatozoide y del
óvulo -cada uno de los cuales se encuentra en su propio gametangio- se
fusionan y producen un cigoto.
Los oomicetes también
pueden reproducirse formando esporas asexuales, cada una de las cuales lleva
dos flagelos.
La mayoría de los oomicetes
son saprobios, es decir, se alimentan de restos de organismos muertos. A pesar
de ello, existen algunas formas parásitas que son patógenas.
Heterótrofos unicelulares
Además de los heterótrofos
multinucleados y multicelulares considerados, el reino Protista incluye un gran
número de heterótrotos unicelulares. Éstos son conocidos
informalmente como protozoos; este grupo está constituido por cinco grupos
principales. Tres de ellos contienen miembros tanto de vida libre como parásitos,
y se distinguen según su modo de locomoción: los zooflagelados
o mastigóforos tienen movimiento flagelar; los ameboides o sarcodinos
emiten seudópodos, y los ciliados o cilióforos tienen movimientos
ciliares. Existen también dos grupos parásitos; los opalínidos,
que tienen movimiento flagelar y los esporozoos, en los cuales casi no existe
movilidad celular.
Los protozoos habitualmente se reproducen asexualmente, por fisión binaria. Muchos también tienen ciclos sexuales que involucran meiosis y fusión de gametas que da lugar a un cigoto diploide (2n). El cigoto frecuentemente adquiere la forma de una cigospora resistente de pared gruesa, especialmente durante los períodos de sequía o frío. Los ciliados, en cambio, experimentan conjugación, en la cual se intercambian núcleos entre las células.

Cubiertas externas, o tecas, características de ciertos grupos de sarcodinos. Arcella dentata .
La teca brillantemente colorada de Arcelladentata consiste en un material proteico
secretado por el organismo.
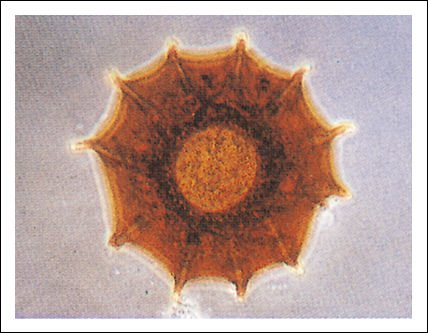
La cubierta de un foraminífero.
Además de las numerosas especies actuales de foraminíferos, hay
aproximadamente 30.000 especies extinguidas, conocidas solamente por sus tecas
fosilizadas.
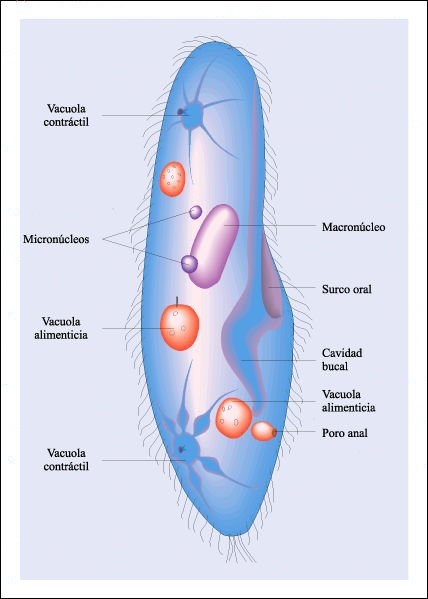
Esquema del ciliado Paramecium.
En la figura anterior se observa un núcleo grande, el macronúcleo
y uno a varios núcleos pequeños, los micronúcleos. Su cuerpo
está completamente cubierto por cilios (estructura 9 + 2) aunque sólo
unos pocos se muestran aquí. Como otros ciliados, Paramecium se alimenta
fundamentalmente de bacterias, de microorganismos de menor tamaño y partículas
de otros materiales. El batir de cilios especializados dirige las partículas
a la cavidad oral donde se forman vacuolas alimenticias que entran al citoplasma.
El alimento es digerido en las vacuolas y la materia no digerida que permanece
en las vacuolas se elimina a través del poro anal. Las vacuolas contráctiles
eliminan el exceso de agua de la célula.
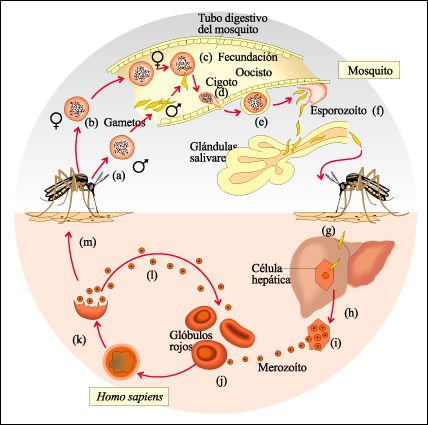
Ciclo de vida de Plasmodium vivax, uno de los esporozoos que causan la malaria en los humanos.
El ciclo comienza a) cuando una hembra de mosquito Anopheles pica a una persona
con malaria y, junto con la sangre, succiona gametas indiferenciadas b) del
esporozoo. En el tracto digestivo del mosquito, las gametas se diferencian,
se unen, c) y forman un cigoto, d). A partir de los cigotos se desarrollan estructuras
multinucleadas llamadas oocistos, e) que, en unos pocos días, se dividen
en miles de células fusiformes muy pequeñas, los esporozoítos,
f). Éstas luego migran a las glándulas salivales del mosquito.
Cuando la hembra pica a otra víctima, g), la infecta con los esporozoítos.
Éstos primero entran a las células hepáticas, h), donde
sufren divisiones múltiples, i). Los productos de estas divisiones (merozoítos)
entran a los glóbulos rojos, j), donde nuevamente se dividen en forma
repetida, k), rompen los glóbulos rojos, 1) a intervalos regulares de
aproximadamente 48 horas; así, provocan episodios febriles recurrentes
que son característicos de esta enfermedad. Después de un período
de reproducción asexual, parte de los merozoítos se transforman
en gametas indiferenciadas (m) y, si son ingeridos por un mosquito en este estadio,
el ciclo comienza nuevamente.
Patrones de comportamiento de
los protistas
Como hemos visto en capítulos
anteriores, incluso los organismos procariotas son capaces de responder a estímulos
ambientales, tales como la luz o sustancias químicas.
Los protistas fotosintéticos,
como Euglena, son muy sensibles a la intensidad lumínica. Nadan hacia
áreas con niveles óptimos de luz y se alejan de las áreas
donde la luz es demasiado brillante. Los no fotosintéticos, como las
amebas, pueden también exhibir un tipo de taxis. En el caso de respuesta
a la luz se denomina fototaxis. Si un seudópodo en avance de una ameba
recibe un haz puntiforme intenso de luz, éste se retrae. Si el cuerpo
entero de la ameba es expuesto a luz brillante, la célula se contrae
repentinamente y es posible que expulse alimento a medio digerir. Sin embargo,
si no puede escapar de la luz, después de una perturbación momentánea
la ameba reanuda sus actividades normales.
Por medio de este tipo de respuesta,
el estímulo llega a ser ignorado y se restablece el patrón de
conducta previa.
Por otra parte, cuando una ameba
percibe algo comestible, una célula de un alga u otro protozoo como ella
que se encuentra en su vecindad, emite un seudópodo cuya forma se ajusta
específicamente a la presa.
Si ésta es pequeña
y quieta, proyecta una fina extensión parecida a una pinza, si se trata
de un ciliado o algo que se mueve activamente, enviará un seudópodo
mucho más grueso.
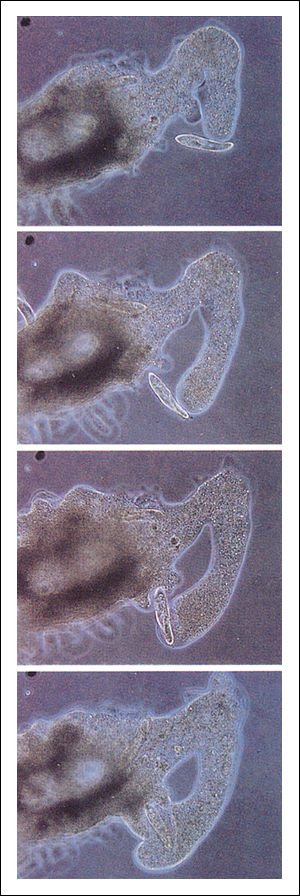
La ameba gigante Chaos chaos capturando una presa.
El estímulo inicial producido por la presa -un Paramecium - induce la
formación de seudópodos en la ameba. La ameba se mueve entonces
hacia el Paramecium. Un gran seudópodo rodea al Paramecium, lo engloba
y forma una vacuola.


