
Capítulo 30.
Las
plantas
Las plantas son organismos fotosintéticos multicelulares adaptados a
la vida terrestre. Entre sus adaptaciones están una cutícula cérea,
poros a través de los cuales intercambian gases, capas protectoras de
células que rodean a las células reproductoras y retención
del esporofito joven dentro del gametofito femenino durante el desarrollo del
embrión.
Todas las plantas parecen haber
surgido de las algas verdes (división Chlorophyta). A diferencia de los
animales que claramente han colonizado la tierra firme repetidas veces a lo
largo de su evolución, la colonización de la tierra por las plantas
parece haber sido un evento único en la historia de la vida.
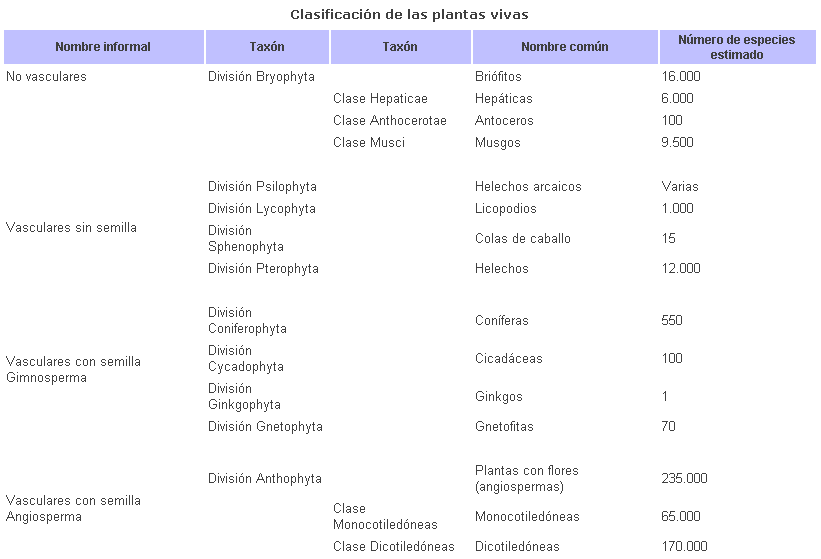
Las plantas modernas se pueden
clasificar en diez divisiones separadas. A partir de un antecesor común,
divergieron dos linajes principales: los briofitos y las plantas vasculares.
La mayoría de los briofitos
carece de tejidos vasculares especializados y todas carecen de hojas verdaderas,
aunque el cuerpo de la planta se diferencia en tejidos fotosintéticos,
de almacenamiento, de alimento y de fijación. Aunque los briofitos parecen
haber cambiado poco en el curso de su historia evolutiva, las plantas vasculares
han sufrido una gran diversificación.
Las principales tendencias que
se observan en la evolución de las plantas vasculares incluyen sistemas
de conducción más eficientes, una reducción progresiva
en el tamaño del gametofito y la aparición de la semilla. Las
nueve divisiones de plantas vasculares pueden agruparse informalmente en las
plantas vasculares sin semillas (divisiones Psilophyta, Lycophyta, Sphenophyta
y Pterophyta) y las plantas con semilla. Las plantas con semilla pueden agruparse
en gimnospermas, o plantas con semillas desnudas (divisiones Coniferophyta,
Cycadophyta, Ginkgophyta y Gnetophyta) y las angiospermas, o plantas que tienen
semillas protegidas que dan flores (división Antophyta).
Entre las plantas vasculares
actuales sin semilla, los helechos (división Pterophyta) son los más
numerosos. Están caracterizados por hojas grandes, a menudo finamente
divididas, llamadas frondes.
Las plantas vasculares con semilla comprenden las gimnospermas y las angiospermas.
Las gimnospermas modernas más numerosas son las coníferas.
Las angiospermas se caracterizan
por la flor y el fruto. Las flores atraen a los polinizadores y los frutos facilitan
la dispersión de las semillas.
Las angiospermas son las plantas
predominantes del paisaje moderno que suministran una diversidad de hábitat
y alimentos para los animales terrestres.
El origen de las plantas superiores
Se cree que el antecesor de
las plantas ha sido un alga verde multicelular. Este alga pertenece a la clase
Charophyceae y es semejante al género moderno Coleochaete que es oogámica.
Al igual que las plantas, las
algas verdes contienen clorofilas a y b y beta-caroteno como pigmentos fotosintéticos,
y acumulan su reserva de alimento en forma de almidón. En las plantas
y las algas verdes, pero no en otros organismos, el almidón se almacena
en plástidos, y no en el citoplasma. Sin embargo, más allá
de estas semejanzas, las algas verdes exhiben una gran diversidad de características,
algunas de las cuales comparten con las plantas y otras no.
Aunque los antecesores inmediatos
de Coleochaete pueden no haber sido los mismos a partir de los cuales evolucionaron
las plantas, habría otro antecesor íntimamente relacionada con
ambos. Varios conjuntos de datos llevaron a esta conclusión. Las células
de Coleochaete, al igual que las de las plantas, tienen celulosa en sus paredes
celulares y contienen peroxisomas en los que se encuentran enzimas claves en
procesos tales como la fotorrespiración, entre otros. Una evidencia adicional
la aporta su patrón de división del citoplasma -citocinesis-.
En casi todos los otros organismos, incluyendo la mayoría de las algas
verdes, el citoplasma se divide por constricción e invaginación
de la membrana celular. En las plantas y en Coleochaete, el citoplasma se divide
por la formación de una placa celular en el ecuador del huso. También
se encuentran similitudes en la estructura y organización de los microtúbulos
y en la estructura de la base de los flagelos de Coleochaete y de las células
vegetales flageladas.
Una característica importante
compartida por todas las plantas, pero ausente en Coleochaete, es una alternancia
de generaciones bien definida. Este tipo de ciclo de vida se encuentra no sólo
en muchas algas verdes multicelulares, sino también en las algas pardas
y en las algas rojas; la evidencia bioquímica indica que se éstos
no son organismos emparentados con las algas verdes. Esto sugiere que la alternancia
de generaciones apareció independientemente en varias ocasiones.
La transición a la tierra
En la época en que el
antecesor inmediato de las plantas se trasladó desde aguas poco profundas
a la tierra firme, aparentemente ya había desarrollado una alternancia
de generaciones heteromórficas bien definida. Después de la transición
a la tierra firme, se fueron seleccionando nuevas adaptaciones en el ciclo de
vida y también en otras características.
Algunas adaptaciones fueron
críticas para el éxito final de las plantas en tierra firma y
deben haber ocurrido temprano en su historia evolutiva, pues la mayoría
de las plantas modernas, aunque sean muy diversas, las comparten. Las células
necesitan agua; de hecho, son 95% de agua. Fue necesario que se reconciliaran
esta exigencia básica de agua en los sistemas biológicos con una
vida en el suelo árido y desnudo de la superficie terrestre. Las plantas
presentan rasgos tales como raíces que las anclan al suelo y que forman
un sistema de captación de agua, sistemas de bombeo, órganos fotosintéticos,
superficies recubiertas contra la desecación y tejidos reforzados.
Estos rasgos son los que permiten que ejemplares como los abetos y tsugas de la siguiente fotografía se eleven decenas de metros sobre la superficie y tengan troncos de varios metros de diámetros.
La transición a la tierra firme fue un evento remarcable e improbable.
Una de estas características,
claramente asociada con la transición a la tierra, es la cutícula
protectora que cubre las superficies aéreas de las plantas y retarda
la pérdida de agua por el cuerpo. La cutícula está formada
por una sustancia cerosa llamada cutina, secretada por las células de
la epidermis. Asociadas con la cutícula y, de hecho, vueltos necesarios
por su existencia, hay "poros" especializados a través de los
cuales pueden realizarse los intercambios de gases necesarios para la fotosíntesis.
Otra adaptación fue el desarrollo de órganos reproductores multicelulares como los gametangios -donde se producen gametas- y los esporangios -donde se producen esporas-. Éstos fueron rodeados por una capa protectora de células estériles. Cuando los gametangios originan gametos femeninos -denominados ovocélulas u oósferas- se llaman arquegonios y cuando originan gametos masculinos o espermatozoides se llaman anteridios.
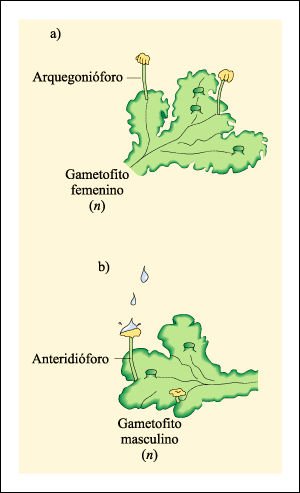
Gametangios multicelulares de la hepática Marchantia, miembro de la división Bryophyta.
a) Gametangios femeninos o arquegonios en varias etapas de desarrollo. Los arquegonios
tienen forma de botella, en cuya base se desarrolla una sola ovocélula.
b) Anteridio en desarrollo en el gametofito masculino. El tejido espermatogénico
originará las gametas masculinas que, cuando estén maduros, nadarán
hasta la ovocélula a través del canal del cuello del arquegonio.
Una adaptación que se relaciona con esto fue la retención de la ovocélula fecundada (el cigoto) dentro del gametangio femenino (el arquegonio) y el desarrollo del embrión dentro de esa estructura. Así, durante las etapas críticas de su desarrollo, el embrión, o esporofito joven, está protegido por los tejidos del gametofito femenino.

Briofitos representativos, las únicas plantas en las cuales el gametofito, que es haploide (n), es la generación dominante y nutricionalmente independiente.
Un gametofito joven de la hepática Marchantia (clase Hepaticae) creciendo
sobre una roca.

Anthoceros (clase Anthocerotae).
Los "cuernos" de este briofito son los esporofitos diploides (2n),
que están unidos a un gametofito con forma de disco.

Musgo politrico (clase Musci) con cápsulas esporíferas.
Las estructuras verdes inferiores del briofito de la figura son los gametofitos.
Los pedicelos y cápsulas no fotosintéticos son los esporofitos.
Según la especie, los esporofitos de los musgos que crecen en regiones
templadas tardan entre 6 y 18 meses en alcanzar la madurez.
Un tiempo después de
la transición a la tierra, las plantas divergieron en al menos dos linajes
separados. El más antiguo, a juzgar por el análisis de la filogenia
basado en la secuencia de la subunidad pequeña del RNA ribosomal dio
lugar a los briofitos, miembros de la división Bryophyta, un grupo que
incluye a las clases hepáticas, antoceros y los musgos modernos.
El otro dio lugar a las plantas
vasculares, el grupo que incluye a todas las restantes plantas terrestres. Una
diferencia principal entre los briofitos y las plantas vasculares es que los
esporofitos de las últimas -conocidos como traqueofitos- tal como su
nombre lo indica, tienen un sistema vascular bien desarrollado que transporta
agua, minerales, azúcares y otros nutrientes a través del cuerpo
de la planta. Los briofitos aparecen por primera vez en el registro fósil
a fines del período Silúrico, hace aproximadamente 400 millones
de años. Estos fósiles antiguos son muy similares a los briofitos
que viven actualmente. Los fósiles más antiguos de las plantas
terrestres -a juzgar por los hallazgos de esporas- son de mediados del período
Ordovícico, hace aproximadamente 470 millones de años.
Así como la aparición de los primeros organismos fotosintéticos tuvo un efecto decisivo en la historia ulterior de la vida, la aparición de las plantas terrestres tuvo importantes consecuencias sobre los flujos de energía y nutrientes entre los ecosistemas terrestres y acuáticos, así como sobre la subsecuente evolución de los animales. Se cree que la aparición de las plantas terrestres fue un factor de gran importancia en la reducción de la concentración atmosférica de dióxido de carbono y -a través de tal reducción- en el descenso de la temperatura sobre la superficie terrestre.
Esquema global de las relaciones evolutivas de los seres vivos.
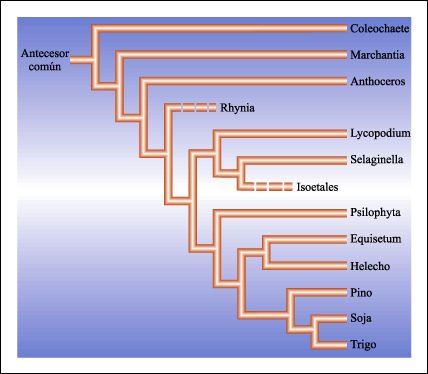
Relaciones filogenéticas simplificadas entre los principales grupos de plantas.
Clasificación de las plantas
Las plantas se pueden clasificar
en dos grandes grupos: los briofitos y las plantas vasculares.
Las hepáticas, antoceros
y musgos de la división Bryophyta son bastante diferentes unos de otros
y hay cierta duda acerca de si representan tres linajes distintos derivados
de la planta ancestral o ramificaciones posteriores de un linaje surgido de
ese antecesor.
Diversas líneas de evidencia
sugieren que cada una de las nueve divisiones de las plantas vasculares es monofilética,
es decir que todos sus miembros descienden de un antecesor común. Las
plantas vasculares se agrupan frecuentemente, por conveniencia, de manera que
pueden o no reflejar relaciones evolutivas. Por ejemplo, estas plantas como
grupo a menudo se conocen como traqueofitos. Pueden ser agrupadas en las que
no tienen semillas (divisiones Psilophyta, Lycophyta, Sphenophyta y Pterophyta)
y en aquellas que tienen semillas. Las plantas con semillas también forman
dos grupos informales, las gimnospermas y las angiospermas. Las gimnospermas
son aquellas que tienen semillas "desnudas", sin protección
(divisiones Coniferophyta, Cycadophyta, Ginkgophyta y Gnetophyta), en tanto
que las angiospermas (de la palabra griega "angio" que significa "vaso"
-literalmente, semilla en un vaso-) son, formalmente hablando, los miembros
de la división Anthophyita, las plantas que dan flores.
División Bryophyta: hepáticas,
antoceros y musgos
Los briofitos carecen de un
sistema radical bien desarrollado y de estructuras altamente especializadas
en el transporte de agua. Por lo tanto, exhiben una complejidad considerablemente
menor que las plantas vasculares. Crecen exitosamente en parajes húmedos
y sombreados, y en ciénagas.
Los briofitos, en general, exhiben
una estructura comparativamente simple y son relativamente pequeños,
habitualmente de menos de 20 centímetros de longitud. Una sola planta
de musgo puede desparramarse en un área considerable, pero la mayoría
de las hepáticas son tan pequeñas que sólo son perceptibles
para un observador agudo. En los ambientes húmedos frecuentados por los
briofitos, las células individuales pueden absorber el agua y los nutrientes
contenidos en el aire o bien incorporados por difusión desde el medio
en que crecen. A semejanza de los líquenes, los briofitos son indicadores
sensibles de la contaminación del aire. Aunque los briofitos no tienen
raíces verdaderas, generalmente se fijan al sustrato por medio de rizoides
que son células individuales alargadas o filamentos celulares. Muchos
briofitos también tienen pequeñas estructuras foliáceas
en las cuales ocurre la fotosíntesis. Estas estructuras carecen de los
tejidos especializados que tienen las hojas "verdaderas" de las plantas
vasculares y tienen sólo el grosor de una o unas pocas capas de células.
Si bien tienen esta lejana similitud, las estructuras foliáceas de los
briofitos y las hojas de las plantas vasculares han evolucionado de modo independiente.
Los briofitos, como todas las otras plantas, tienen un ciclo de vida con alternancia de generaciones. Sin embargo, en contraste con las plantas vasculares, los briofitos se caracterizan por poseer un gametofito haploide que, en general, es de mayor tamaño que el esporofito diploide.
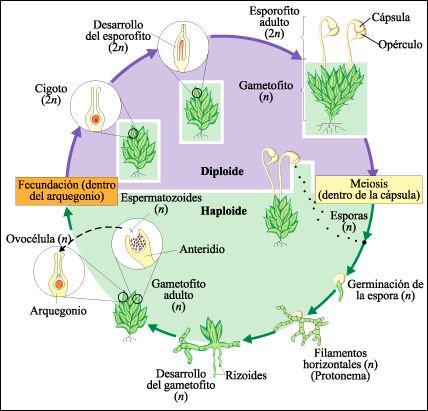
El ciclo de vida de un musgo.
El ciclo de vida de un musgo comienza con la liberación de esporas de
la cápsula, que se abre cuando se expulsa una pequeña tapa denominada
opérculo (arriba derecha). La espora germina y da lugar a un protonema
filamentoso ramificado, a partir del cual se desarrolla un gametofito foliáceo.
Los espermatozoides, que son expulsados del anteridio maduro, son atraídos
al arquegonio donde uno de ellos se fusiona con la ovocélula y produce
el cigoto. El cigoto se divide por mitosis y forma el esporofito. Al mismo tiempo,
la base del arquegonio se divide y forma la caliptra protectora. El esporofito
maduro consiste en una cápsula, que puede estar sostenida por un pedicelo
-que es también parte del esporofito- y un pie. La meiosis ocurre dentro
de la cápsula y da como resultado la formación de esporas haploides.
En este musgo, los gametofitos llevan tanto anteridios como arquegonios. En
otras especies, un sólo gametofito puede llevar anteridios o arquegonios,
pero no ambos.

Gametofito de la hepática Marchantia.
Las copas gemíferas con forma de cuenco, visibles en este gametofito
de la hepática Marchantia, contienen cuerpos diminutos, los propágulos
que son esparcidos por la lluvia y crecen en la vecindad de la planta madre.
La reproducción asexual por fragmentación o por propágulos
es común entre las hepáticas. En Marchantia, los anteridios y
arquegonios elevados se forman en plantas diferentes: el gametofito masculino
b) y el femenino. c) El cigoto, formado en el arquegonio, desarrolla un esporofito,
que permanece unido al gametofito femenino. Las copas gemíferas se desarrollan
en ambos tipos de gametofito.
Las plantas vasculares: introducción
Rhynia major, actualmente extinguida,
es un ejemplo de las plantas vasculares más antiguas que se conocen y
su existencia se remonta a unos 400 millones de años atrás. Su
aspecto parece mucho más primitivo que el de un briofito. Sin embargo,
difería de los briofitos en un punto importante: dentro de su tallo había
un cilindro central de tejido vascular, especializado en la conducción
de agua y sustancias disueltas hacia la parte superior del cuerpo.
Comenzando con una planta vascular
muy simple, tal como Rhynia, es posible seguir algunas tendencias evolutivas
principales, así como diversas innovaciones fundamentales. Una innovación
temprana fue la raíz, estructura especializada en la fijación
de la planta y la absorción de agua y nutrientes esenciales. Otra fue
la hoja, estructura especializada en la fotosíntesis.
Rhynia major carecía de hojas y de raíces. Sus tallos aéreos, que eran fotosintéticos, estaban unidos a un tallo subterráneo. o rizoma. Los tallos aéreos estaban cubiertos con una cutícula y contenían estomas. Las estructuras oscuras en los ápices de los tallos son esporangios, que aparentemente liberaban sus esporas al escindirse longitudinalmente.
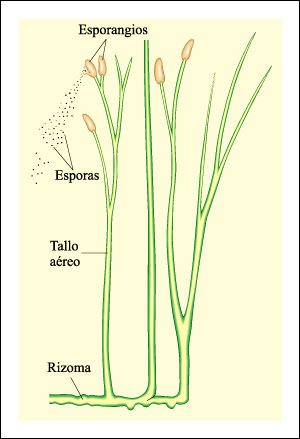
Rhynia major, una de las plantas vasculares más primitivas que se conocen.
Uno de los hechos más sorprendentes en la evolución de las plantas
ha sido el desarrollo de sistemas conductores cada vez más eficientes
que comunican las dos porciones del cuerpo de la planta, el vástago (que
comprende el tallo y las hojas) y la raíz.
El sistema conductor en las
plantas vasculares modernas consta de dos tejidos diferentes: el xilema, que
transporta agua e iones desde las raíces a las hojas y el floema, que
lleva sacarosa y otros productos de la fotosíntesis disueltos, desde
las hojas a las células no fotosintéticas de la planta. Si bien
el transporte de estas sustancias es de vital importancia, el floema también
es empleado para el transporte a larga distancia -entre distintas localidades
de la planta- de muchas otras sustancias. De igual modo, el fluido xilemático
no está compuesto sólo de agua e iones, sino que contiene, con
frecuencia, sustancias orgánicas disueltas.
Los elementos conductores del
xilema son las llamadas traqueidas y los vasos. Los elementos conductores del
floema son las conocidas como células cribosas. Asociadas a estas células
se encuentran las llamadas células acompañantes. En los tallos,
los cordones longitudinales del xilema y del floema corren contiguos, ya sea
en haces vasculares o dispuestos en dos capas concéntricas (cilindros)
en las cuales uno de los tejidos (típicamente el floema) está
por fuera del otro.
Con el desarrollo de raíces,
hojas y sistemas conductores eficientes, las plantas "resolvieron"
efectivamente los problemas más básicos con los que se enfrentaban
los organismos multicelulares fotosintéticos en tierra: adquirir abastecimientos
adecuados de agua y nutrientes, y distribuirlos entre todas las células
que constituyen el organismo.
Otra tendencia pronunciada que
se observa en la evolución de las plantas es la reducción en el
tamaño del gametofito. En todas las plantas vasculares y, a diferencia
de lo que hemos visto en los briofitos, el gametofito es más pequeño
que el esporofito. Sin embargo, en los representantes contemporáneos
de las plantas vasculares más primitivas, el gametofito está separado
y es nutricionalmente independiente del esporofito. En los grupos que han evolucionado
más recientemente -las gimnospermas y las angiospermas- el gametofito
se ha reducido a un tamaño microscópico y a una condición
de extrema dependencia respecto del esporofito.
Relacionada también con
el ciclo reproductor, hay una tendencia hacia la heterosporia, es decir, la
producción de dos tipos de esporas. Las plantas vasculares más
primitivas producían sólo un tipo de espora (homosporia) en un
tipo de esporangio. A1 germinar, estas esporas típicamente producen gametofitos
en los cuales se forman tanto anteridios como arquegonios. En las plantas que
son heterósporas, se desarrollan un gametofito que lleva los arquegonios
y otro que lleva los anteridios. A medida que los gametofitos se fueron reduciendo
de tamaño, los arquegonios y los anteridios también disminuyeron
de tamaño, hasta desaparecer completamente en las angiospermas. En las
angiospermas, así como en las gimnospermas, el gametofito masculino recibe
el nombre de polen.
El gametofito femenino, en tanto,
produce células haploides contenidas en una estructura denominada óvulo.
El óvulo, en las plantas angiospermas, consta del gametofito femenino
y el tegumento (2n) que lo recubre. El óvulo fecundado o semilla es una
de las innovaciones más importantes que contribuyen a explicar el enorme
éxito de las plantas vasculares en tierra firme. La semilla es una estructura
compleja que contiene al esporofito joven, o embrión, rodeado de una
cubierta externa protectora, la cubierta seminal. Esta cubierta, que deriva
de tejidos del esporofito materno, protege al embrión mientras éste
permanece latente, a veces durante muchos años, hasta que las condiciones
sean favorables para su germinación. Las semillas más tempranas
que se conocen se fosilizaron en depósitos del Devónico superior,
hace aproximadamente 360 millones de años.
Las plantas vasculares sin semillas
Hay cuatro divisiones de plantas
vasculares sin semillas que tienen representantes vivos: las Psilophyta (helechos
arcaicos), las Lycophyta (licopodios), las Sphenophyta (colas de caballo) y
las Pterophyta (helechos), el grupo más grande. Muchas de ellas ya estaban
presentes en el perído Carbonífero.
De acuerdo con el testimonio
de los fósiles, los helechos aparecieron hace unos 350 millones de años,
y todavía son relativamente abundantes. La mayoría de las 12.000
especies vivientes se encuentran en los trópicos, pero muchas existen
también en regiones templadas y aun áridas. Dado que sus espermatozoides
son flagelados y necesitan de agua libre para la fecundación, las especies
que viven en regiones áridas explotan la aparición estacional
de agua para la reproducción sexual.
Los tallos de los helechos no
son habitualmente tan complejos como los de las gimnospermas y angiospermas,
y a menudo se reducen a un rizoma. Aunque los helechos no poseen crecimiento
secundario, es decir, el tipo de crecimiento que da como resultado un aumento
de la circunferencia y la formación de corteza y tejido leñoso,
algunos alcanzan una gran altura.
Las hojas o frondas de los helechos frecuentemente están finamente divididas en folíolos o pinnas. Caracterizadas por una alta relación superficie a volumen, estas hojas divididas, muy desplegadas, son colectores de luz muy eficientes. Los helechos están bien adaptados a crecer en las condiciones de luz difusa que impera en el suelo de los bosques. Los esporangios comúnmente se disponen en la superficie inferior de las hojas o, a veces, en hojas especializadas. Las hojas que traen esporangios se llaman esporofilos. Los esporofilos pueden asemejarse a las otras hojas verdes de la planta o pueden ser tallos no fotosintéticos (hojas modificadas). Los esporangios de los helechos comúnmente se presentan en pequeños grupos conocidos como soros.

Representantes actuales de las cuatro divisiones de plantas vasculares sin semillas.
a) El helecho Psilotum, uno de los dos géneros vivos de la división
Psilophyta. Las estructuras bulbosas son los esporangios, que aparecen en grupos
fusionados de a tres. Estos helechos arcaicos son únicos entre las plantas
vasculares actuales por carecer de hojas y raíces. Sin embargo, si se
mira con detenimiento, se distinguen unas pequeñas emergencias en forma
de escamas debajo de los esporangios. b) Los licopodios, helechos del género
Lycopodium, son los miembros más comunes de la división Lycophyta.
En este género, los esporangios son llevados en hojas especializadas,
los esporofilos, que se agrupan formando un cono en el ápice (parte superior)
de las ramas, como se muestra en este Lycopodium clavatum. Las esporas céreas,
transportadas por el viento, originan pequeños gametofitos subterráneos
independientes. Los que son biflagelados, nadan hasta el arquegonio, donde se
produce la fecundación y se desarrolla el esporofito joven, o embrión.
c) Los colas de caballo, división Sphenophyta, de los cuales hay un sólo
género vivo (Equisetum ) se reconocen fácilmente por sus tallos
articulados y finamente estriados, que contienen sílice. En cada nudo
hay un círculo de hojas pequeñas, en forma de escamas. Las estructuras
que llevan esporas se agrupan en un cono en el ápice del tallo. Los gametofitos
son independientes y los espermatozoides son espiralados y poseen numerosos
flagelos. d) Un helecho real. Los helechos, división Pterophyta, son
las plantas sin semillas más abundantes.
En los helechos, como en todas las plantas vasculares vivientes, la generación dominante es el esporofito.
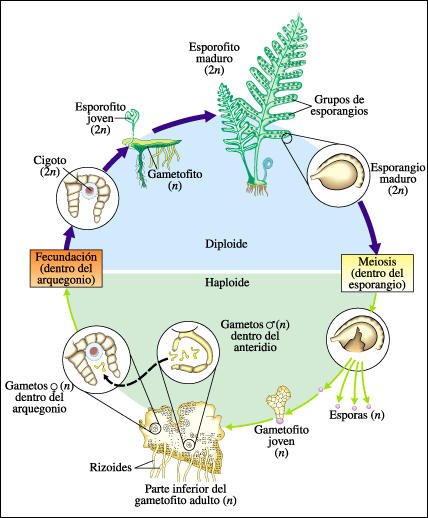
Ciclo de vida de un helecho.
En los esporangios (a la derecha), se producen, por meiosis, las esporas haploides,
que luego son liberadas. De las esporas se desarrollan gametofitos haploides.
En muchas especies, los gametofitos tienen sólo unas pocas capas de células
y adoptan una forma similar a un corazón, como se muestra aquí
(parte inferior). De la superficie inferior del gametofito aparecen filamentos,
los rizoides, que penetran en el suelo. En la superficie inferior del gametofito
hay arquegonios, estructuras con forma de botella, que contienen las gametos
femeninos y los anteridios, que contienen a los gametos masculinos. Cuando los
gametos masculinos maduran y hay un aporte adecuado de agua, los anteridios
se rompen y los gametos masculinos, que tienen numerosos flagelos, nadan hasta
los arquegonios y fecundan a los gametos femeninos. Del cigoto se desarrolla
el esporofito diploide (2n), que crece del arquegonio, contenido en el gametofito.
Después que el joven esporofito se arraigó en el suelo, el gametofito
se desintegra. El esporofito madura, desarrolla esporangios, en los cuales ocurre
la meiosis y así comienza nuevamente el ciclo.
Plantas con semillas
Las plantas con semillas existían
ya cerca del final del período Carbonífero. Por entonces, y de
acuerdo con el registro fósil, la exuberante vegetación estaba
dominada por helechos y licopodios arborescentes de gran tamaño.
Durante el período Pérmico, las gimnospermas se diversificaron. Cuatro grupos de gimnospermas tienen representantes vivos: tres divisiones pequeñas -Cycadophyta, Ginkgophyta y Gnetophyta- y una división grande y familiar para todos nosotros -Coniferophyta-. Las coníferas ("portadoras de conos") incluyen a los pinos, abetos, piceas, Tsuga del Canadá, juníperos, alerces y araucarias de Argentina y Chile, así como las secuoyas gigantes de California y Oregon.

Gimnospermas representativas.
Encephalartos Woodii, una cicadácea (división Cycadophyta) procedente
de África. La mayoría de las cicas son raras. Esta especie, que
se encuentra en peligro de extinción, se ha salvado debido a que muchos
rebrotes de una planta progenitora se encuentran a buen resguardo en varios
jardines botánicos del mundo.

Gingko biloba
Hojas y semillas carnosas del Gingko biloba la única especie sobreviviente
de las Gingkophyta, un linaje que data del Paleozoico superior. Gingko es especialmente
resistente a la contaminación del aire y comúnmente se cultiva
en parques urbanos y a lo largo de las calles de la ciudad. La cubierta carnosa
de semillas tiene un olor pútrido semejante a la manteca rancia. Sin
embargo, la "almendra interna de la semilla", que tiene un sabor a
pescado, es un manjar muy apreciado en el Oriente.

Pino Ponderosa
Rama de conífera, del pino Ponderosa (también llamado pino amarillo
del oeste), con un cono femenino. Cuando el cono madura, se abre y libera sus
semillas aladas (en la figura se observan dos semillas que han quedado atrapadas
entre las escamas). Al igual que otros pinos, el Ponderosa tiene hojas aciculares
-en forma de aguja- flexibles, que se mantienen juntas en un haz. El pino Ponderosa,
uno de los principales árboles de los bosques de las Rocosas, desde Canadá
a México, es un pilar para la industria maderera del noroeste de América
del Norte y otras partes del mundo.

Welwitschia mirabilis
Una gran planta productora de
semillas, Welwitschia mirabilis, un gnetófito (división Gnetophyta),
que crece en el desierto de Namibia de África meridional. La Welwitschia
produce solamente dos hojas adultas, que continúan su crecimiento durante
toda la vida de la planta. A medida que el crecimiento continúa, las
hojas se rompen en los extremos y se escinden a lo largo. Así, las plantas
más viejas parecen tener numerosas hojas.
La semilla es una estructura
protectora por medio de la cual los embriones pueden dispersarse y permanecer
latentes hasta que las condiciones se tornen favorables para su supervivencia.
Así, sus funciones se asemejan a las esporas de las bacterias o a los
cigotos resistentes de las algas de agua dulce. Si bien existe esta superficial
similitud, la estructura de las semillas es mucho más compleja. Una semilla
incluye el embrión (el esporofito latente, joven), una reserva de tejido
nutritivo y una cubierta protectora externa.
En las plantas con semillas,
la generación del gametofito se reduce aun más y depende totalmente
del esporofito.
Todas las gimnospermas son heterósporas y producen dos tipos diferentes de esporas en dos tipos diferentes de esporangios. Las esporas que originan los gametofitos masculinos se conocen como micrósporas y se forman en estructuras conocidas como microsporangios. Las esporas a partir de las cuales se desarrollan los gametofitos femeninos, se conocen como megaspora y se forman en los megasporangios. Un megasporangio contiene una sola célula madre de la megaspora, que origina, por meiosis, a una megaspora, y está rodeada por una o dos capas de tejido, el tegumento.

Ciclo de vida del pino.
Las estructuras reproductoras son los conos, dentro de los cuales se forman
las esporas sobre las escamas. Las microsporas se desarrollan a partir de las
células madre de las microsporas y las megasporas, a partir de las células
madre de las megasporas. Las microsporas desarrollan granos de polen, que son
gametofitos masculinos inmaduros. Dentro de los óvulos, las megasporas
desarrollan un gametofito femenino; cada gametofito femenino contiene varios
arquegonios, cada uno con una ovocélula. Aunque más de una ovocélula
pueda ser fecundada, habitualmente sólo se desarrolla completamente un
embrión en cada gametofito femenino. Los gametos masculinos inmóviles
son llevados al arquegonio por el tubo de polen, y la ovocélula es fecundada.
Después de la fecundación, el óvulo madura formando la
semilla; la semilla consiste en el esporofito embrionario, que rodea al tejido
nutritivo del gametofito femenino y una cubierta externa derivada de las capas
protectoras (tegumento) del óvulo. Cuando la semilla madura, el cono
se abre y libera las semillas aladas que germinan produciendo la plántula.
Ambos tipos de conos se desarrollan en el mismo esporofito maduro.

Conos masculinos del pino de Escocia ( Pinus sylvestris), diseminando su polen.
Los granos de polen son gametofitos masculinos inmaduros que completan su maduración
cuando alcanzan los óvulos incluidos en los conos femeninos. Allí
producen tubos de polen que transportan los gametos masculinos inmóviles
a las ovocélulas.

Cono femenino del pino de Escocia.
Los gametofitos femeninos se desarrollan en óvulos en la base de una
escama del cono, y las ovocélulas son fecundadas allí. Cada escama
contiene dos óvulos. Cuando las semillas maduran, caen del cono.
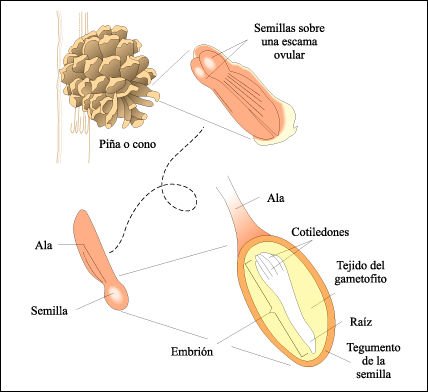
Semilla de pino.
Las capas externas, es decir, el tegumento del óvulo se han endurecido
y forman una cubierta seminal que encierra al gametofito femenino y al embrión,
que ahora consiste en una raíz embrionaria y varias hojas embrionarias,
los cotiledones. Cuando la semilla germina, la raíz emergerá de
la cubierta de la semilla y penetrará en el suelo. Cuando la raíz
absorbe agua, los cotiledones fuertemente compactados se alargarán y
se hincharán con la humedad, surgiendo por encima de la superficie del
suelo sobre el tallo que se alarga y se desprenderán de la cubierta de
la semilla. Durante este período, los cotiledones absorben nutrientes
almacenados en el tejido del gametofito, que son esenciales para el desarrollo
del embrión en una plántula.
Se cree que las angiospermas
-plantas con semillas encerradas y protegidas- evolucionaron a partir de un
grupo actualmente extinguido de gimnospermas. Aparecieron en el registro fósil
§ en abundancia durante el período Cretácico, hace unos 120
millones de años, cuando los dinosaurios estaban en su apogeo.
El momento en que se originaron
las angiospermas es aún objeto de debate.
Las angiospermas tienen dos
estructuras nuevas interrelacionadas, que las distinguen de todo el resto de
las plantas: la flor y el fruto. Ambas estructuras están relacionadas
con la reproducción y dispersión de las plantas.
Se conocen aproximadamente 235.000
especies de angiospermas. Dominan las regiones tropicales y templadas del mundo,
ocupando más del 90% de la superficie vegetal de la Tierra. En la actualidad,
las angiospermas incluyen no sólo a las plantas con flores conspicuas,
sino también a los grandes árboles de madera dura, a todos los
frutales, hortalizas, hierbas, y a los granos y forrajes que son componentes
básicos de la dieta humana y la base de la economía agrícola
de todo el mundo. Estas plantas tremendamente diversas se clasifican en dos
grandes grupos: la clase de las monocotiledóneas y la clase de las dicotiledóneas.
Entre las monocotiledóneas se encuentran plantas tan familiares como
los pastos (gramíneas), lirios, iris, orquídeas, espadañas
o totoras, y palmeras. Las dicotiledóneas incluyen muchas de las hierbas,
casi todos los arbustos y árboles (excepto las coníferas) y muchas
otras plantas.

Las flores son estructuras reproductivas especializadas en las que se lleva a cabo la reproducción sexual. En las flores se forman las semillas y, a partir de éstas, se desarrollan los frutos
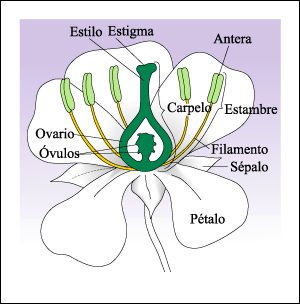
Estructura de una flor.
Algunas flores tienen solamente estructuras masculinas, otras solamente femeninas;
se dice que tales flores son imperfectas. Una flor que posee estambres y carpelo,
como ésta, se conoce como flor perfecta. Los pétalos y los sépalos,
al igual que los estambres y el carpelo, son hojas modificadas.
Los granos de polen producidos en las anteras son usualmente transportados al estigma de otra flor, donde germinan, desarrollando tubos de polen que crecen a través del estilo hacia el óvulo.
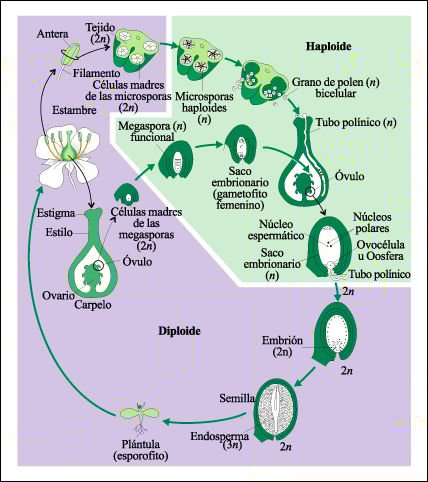
Ciclo de vida de una angiosperma.
Durante el ciclo de vida de una angiosperma, dentro de la antera de la flor,
las células madres de las microsporas se dividen meióticamente
originando, cada una, cuatro microsporas haploides. El núcleo de cada
microspora se divide luego mitóticamente y la microspora desarrolla un
grano de polen bicelular, que es un gametofito masculino inmaduro. Una de las
células se divide posteriormente otra vez, habitualmente después
del desarrollo del tubo polínico, dando como resultado tres células
haploides por grano de polen: dos gametos masculinos inmóviles y la célula
generadora del tubo polínico.
Dentro del óvulo, una
célula madre de la megaspora se divide meióticamente y forman
cuatro megasporas haploides. Tres de las megasporas se desintegran; la cuarta
se divide mitóticamente, da lugar a el saco embrionario -el gametofito
femenino- que consiste en siete células con un total de ocho núcleos
haploides (la célula central grande contiene dos núcleos, los
núcleos polares). Una de las células más pequeñas,
que contiene un solo núcleo haploide es la ovocélula. El polen
germina sobre el estigma, produciendo un tubo de polen que crece a través
del estilo hasta el ovario. El tubo de polen en crecimiento penetra en el óvulo
a través de una pequeña abertura conocida como micrópilo.
Los dos gametos masculinos inmóviles pasan a través del tubo al
saco embrionario; el núcleo de un gameto masculino fecunda a la ovocélula.
El otro se fusiona con los núcleos polares, formando una célula
triploide (3n) que se desarrolla en un tejido nutritivo, el endosperma. El embrión
pasa por sus primeras etapas de desarrollo mientras se encuentra aún
dentro del ovario de la flor, el ovario mismo madura y se transforma en fruto.
La semilla, liberada del esporofito materno en estado latente, germina finalmente
formando una plántula.
Las principales tendencias en
la evolución de las flores incluyen la reducción y fusión
de las piezas florales, un cambio en la posición del ovario con relación
con las otras partes de la flor hacia una posición más protegida
(inferior) y un cambio de la simetría radial a bilateral. Además
de la flor y el fruto, un tercer factor en el éxito de las angiospermas
fue la producción de sustancias químicas de mal sabor o tóxicas
que desalientan las depredaciones por animales forrajeros.
El papel de las plantas
Las únicas formas de
vida sobre la Tierra que no dependen de las plantas para su existencia son unos
pocos tipos de procariotas y protistas autótrofos. Para todos los otros
organismos terrestres, el cloroplasto de la célula vegetal es el "ojo
de la aguja" a través del cual se canaliza la energía solar
a la biosfera. Aun aquellos animales que comen sólo a otros animales,
los carnívoros, no podrían existir si su presa, o la presa de
sus presas, no se hubiera nutrido de plantas. Además, las plantas son
los canales mediante los cuales muchas de las sustancias inorgánicas
simples, esenciales para la vida, entran en la biosfera. El carbono, en forma
de dióxido de carbono, es tomado de la atmósfera e incorporado
a compuestos orgánicos mediante la fotosíntesis.
Los elementos tales como el
nitrógeno y el azufre son tomados del suelo en forma de compuestos inorgánicos
simples e incorporados a proteínas, vitaminas y otros compuestos orgánicos
esenciales dentro de las células vegetales verdes. Los animales no pueden
fabricar estos compuestos orgánicos a partir de materiales inorgánicos
y, por lo tanto, dependen enteramente de las plantas para obtener estos compuestos,
así como para su abastecimiento de energía.
Fue solamente después
de que las plantas hubiesen invadido exitosamente la Tierra, que los miembros
de varios diferentes grupos de animales pudieron, a su vez, llevar a cabo sus
propias invasiones de este vasto nuevo ambiente. Tanto los invertebrados como
los vertebrados experimentaron una gran diversificación cuando aprovecharon
la variedad de hábitat y reservas alimenticias puestas a su disposición
por las plantas.


