
Capítulo 33.
El reino
animal III: los artrópodos
Los artrópodos constituyen el phylum animal más grande, tanto
en número de especies como de individuos. Los artrópodos se caracterizan
por el tipo de apéndices, por su segmentación y exoesqueleto y
por sus órganos sensoriales. En la mayoría de los grupos, los
segmentos se combinan formando una cabeza, un tórax (en algunas ocasiones
fusionado con la cabeza formando el cefalotórax) y un abdomen.
Existen tres grupos principales
de artrópodos: los quelicerados, los mandibulados acuáticos y
los mandibulados terrestres.
Los mandibulados se caracterizan
por poseer quelíceros y pedipalpos; los mandibulados acuáticos,
con dos pares de antenas, un par de mandíbulas y dos pares de maxilas,
y los mandibulados terrestres, con un par de antenas, un par de mandíbulas
y un par de maxilas. Los quelicerados incluyen a los cangrejos cacerola o límulos
(clase Merostomata); las arañas de mar (clase Pycnogonida) y las arañas,
escorpiones y ácaros -entre los cuales están las garrapatas- (clase
Arachnida). Los mandibulados acuáticos, algunos de los cuales viven actualmente
en ambientes terrestres húmedos, pertenecen todos a la clase Crustacea
e incluyen a animales tan comunes como los bogavantes, cangrejos de río,
camarones y percebes. Los mandibulados terrestres incluyen cuatro clases relativamente
pequeñas (Chilopoda, Diplopoda, Pauropoda y Simphyla) y la clase más
grande del reino animal, Insecta, con aproximadamente un millón de especies.
Los insectos son los únicos invertebrados capaces de volar.
Entre los factores que contribuyeron
al extraordinario éxito de los artrópodos están su exoesqueleto,
su tamaño generalmente pequeño y su gran especialización,
tanto en la dieta como en el hábitat. Factores adicionales del éxito
son la capacidad de volar y la metamorfosis completa, que permite un mayor refinamiento
de las adaptaciones para la alimentación, la reproducción y la
dispersión, así como la competencia reducida entre los adultos
y las formas inmaduras.
El sistema nervioso presenta
órganos sensoriales diversos y también ha sido importante en el
éxito de los artrópodos. Entre los receptores sensoriales más
importantes están el ojo compuesto, los receptores táctiles, los
propioceptores y los órganos timpánicos. Los artrópodos
se comunican con miembros de la misma especie mediante el sonido y también
por feromonas.
Características de los
artrópodos
La característica que
define a los artrópodos son sus apéndices articulados. En los
miembros del phylum que tienen el número de apéndices reducido
éstos están, en general, más especializados y son más
eficientes. Particularmente entre los insectos y crustáceos, estos apéndices
especializados incluyen, no sólo patas que usan para caminar, sino también
un conjunto de instrumentos adaptados a las más diversas funciones: mandíbulas,
branquias, lenguas, ovipositores, tubos chupadores, garras, antenas, remos y
pinzas.
Todos los artrópodos están segmentados, una característica
que sugiere fuertemente un antecesor común con los anélidos. Sin
embargo, en el curso de la evolución de los artrópodos, se observa
un cuerpo más corto y con menos segmentos; el número de segmentos
ha quedado fijo y son muy especializados.
Esquema global de las relaciones evolutivas de los seres vivos.

Probable filogenia de los artrópodos.
En muchos artrópodos,
la tagmosis ha progresado mucho más que en los anélidos poliquetos,
pues los segmentos se han fusionado y forman distintas regiones corporales:
una cabeza, un tórax (en algunas ocasiones fusionado con la cabeza para
formar un cefalotórax) y un abdomen. Pero, el patrón básico
de segmentación a menudo se ve claramente en las etapas inmaduras (como
en el caso de la oruga) y puede ser descubierto en el adulto por el examen de
los apéndices, la musculatura y el sistema nervioso.
En algún momento después que la aparición del linaje que condujo a los artrópodos divergió del que condujo a los anélidos, ocurrieron otras diversificaciones importantes. Estas ramificaciones dieron lugar a tres tipos principales de artrópodos: los quelicerados, los mandibulados acuáticos y los mandibulados terrestres. Las diferencias conspicuas que muestran los apéndices de estos tres tipos pueden ser distinguidas claramente, incluso por un ojo inexperto. Tanto en los mandibulados acuáticos (clase Crustácea) como en los mandibulados terrestres (clase Insecta y cuatro clases más pequeñas), los apéndices más anteriores consisten en uno o dos pares de antenas seguidas por un par de mandíbulas y uno o dos pares de maxilas.
Las diferencias en el desarrollo
y la estructura de las mandíbulas sugiere que evolucionaron independientemente
en los dos grupos mencionados, y que no son reflejo de un antecesor mandibulado
común. Los quelicerados, que incluyen la clase Merostomata (límulos),
la clase Pycnogonida (arañas de mar) y la clase Arachnida (arañas,
ácaros, escorpiones y sus parientes), no tienen ni antenas ni mandíbulas.
Su primer par de apéndices son quelíceros, que toman la forma
de pinzas o colmillos.
A pesar del enorme número
de artrópodos y de su rica diversidad, hay un número de características
que comparten todos los miembros de este phylum.
La característica más
notable de todos los artrópodos es su exoesqueleto articulado. Este exoesqueleto
-o cutícula- es secretado por la epidermis subyacente y está adherido
a ella. Está constituido por una capa externa, frecuentemente cérea,
compuesta de lípidos y lipoproteínas; una capa media endurecida
y una capa interna flexible, compuestas ambas principalmente por quitina y otras
proteínas. El exoesqueleto sino que se extiende interiormente, formando
la primera porción de ambos extremos del tubo digestivo. En los insectos
también forma las tráqueas. La cutícula permite la protección
contra los depredadores y es, en general, impermeable. Interviene en la trituración
de alimentos en el intestino anterior y forma además las lentes del ojo
de los artrópodos.
Los músculos están
insertos en diferentes porciones del exoesqueleto, del mismo modo que se insertan
en los diversos huesos del endoesqueleto de los vertebrados. Cuando los músculos
se contraen, el exoesqueleto se mueve al nivel de sus articulaciones.
El exoesqueleto, sin embargo,
tiene ciertas desventajas. No crece y, por tal motivo, debe ser descartado y
vuelto a formar varias veces a medida que el animal crece y se desarrolla. El
proceso que lleva a la formación de una nueva cutícula para despojarse
de la vieja se conoce como muda.La muda culmina en la ecdisis, cuando el animal,
al tragar agua o aire se hincha y rasga el exoesqueleto viejo debilitado. Luego
de liberarse de la cutícula vieja, se hincha; así estira la nueva
cutícula y aumenta de tamaño corporal. El animal recién
emergido es particularmente vulnerable a los depredadores. La muda es un proceso
costoso, en función del gasto metabólico que implica.
Los artrópodos se caracterizan por tener un sistema circulatorio abierto y un sistema nervioso que consiste en una serie de ganglios, un par por segmento, interconectados por un cordón nervioso ventral doble. Las tráqueas (tubos respiratorios tapizados por cutícula), las branquias laminares, los pulmones laminares y los túbulos de Malpighi (conductos excretores que llevan al intestino posterior) se encuentran casi exclusivamente en los artrópodos.
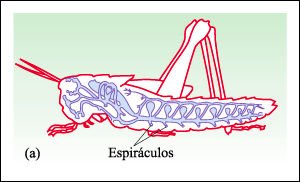
El sistema respiratorio traqueal, ejemplificado en un saltamontes.
La respiración se realiza por medio de un sistema de tubos internos (llamados
tráqueas), que se encuentra casi exclusivamente entre los artrópodos
terrestres. Las tráqueas habitualmente están ramificadas, como
las que se encuentran aquí, y se abren al exterior por espiráculos
que pueden cerrarse y así contribuir a conservar el agua. Los tubos están
formados por cutícula y anillos espirales de cutícula engrosada,
que los mantienen abiertos. Dado que entrega oxígeno directamente a las
células, el sistema de tráqueas es uno de los sistemas respiratorios
más eficientes del reino animal. Sin embargo, a medida que el animal
aumenta de tamaño su eficiencia disminuye

Los túbulos de Malpighi.
Los túbulos de Malpighi representan otra característica casi exclusiva
de los artrópodos aunque, al igual que las tráqueas, no se encuentra
en todas las clases. Estos túbulos colectan agua y desechos nitrogenados
del hemocele y los vacían en el intestino. Los desechos (en forma de
ácido úrico o guanina) son excretados junto con las heces a través
del ano.

El sistema nervioso de los artrópodos, ejemplificado en una abeja.
El cerebro consiste en tres pares fusionados de ganglios dorsales situados en
la cabeza; el extremo anterior está constituido por una doble cadena
de ganglios ventrales. Los ganglios están interconectados por dos haces
de fibras nerviosas que corren a lo largo de la superficie ventral. A raíz
de la naturaleza segmentaria del sistema nervioso del artrópodo, muchas
funciones son controladas a nivel local y varias especies pueden desempeñar
sus actividades normales después de haberse eliminado el cerebro.
Subdivisiones del phylum artropodos
Los artrópodos quelicerados
carecen de antenas y de mandíbulas. El primer par de apéndices,
los quelíceros, que forman pinzas o son agudos y semejantes a colmillos,
se usan para morder a la presa. El segundo par son los pedipalpos, que pueden
tener pinzas, estar modificados como patas locomotoras, o servir como órganos
sensoriales o reproductores. Detrás de los pedipalpos, hay una serie
de patas locomotoras articuladas.
Los segmentos del cuerpo se
encuentran fusionado en dos tagmas: un cefalotórax anterior (o prosoma)
y un abdomen posterior (u opistosoma). Éste no es segmentado en la mayoría
de los quelicerados, pero es conspicuamente segmentado en los escorpiones. Excepto
algunos ácaros, todos los quelicerados son carnívoros y los sexos
casi siempre están separados. Todos los quelicerados restantes pertenecen
a la clase Arachnida.
Los arácnidos, que incluyen
arañas, ácaros (como las garrapatas) y escorpiones, entre otros,
son casi todos terrestres, excepto unas pocas especies que han regresado al
agua. Los arácnidos tienen cuatro pares de patas locomotoras y sus quelíceros
y pedipalpos suelen estar altamente especializados. En las arañas, los
conductos de un par de glándulas ponzoñosas se vacían a
través de los quelíceros, que son afilados y puntiagudos; con
ellos pueden morder y paralizar a la presa.
Los escorpiones usan sus pedipalpos
para tomar y desmenuzar el alimento. Las arañas macho también
usan los pedipalpos para transferir el semen a la hembra.
Las arañas, como la mayoría
de los arácnidos, viven de una dieta completamente líquida. Todas
son predadoras. La presa es mordida y frecuentemente paralizada por los quelíceros.
Luego, las enzimas del intestino medio se vierten sobre los tejidos desgarrados
para producir un caldo parcialmente digerido. Los tejidos licuados de la presa
son bombeados al estómago y luego al intestino, donde se completa la
digestión y se absorben los nutrientes.
Los arácnidos respiran
por medio de tráqueas o pulmones laminares; ambas estructuras pueden
coexistir en un mismo individuo. Los pulmones laminares son sacos que se originan
de una invaginación del tegumento; la pared anterior se pliega formando
una serie de placas en forma de hojas, entre las cuales circula el aire que
entra y sale por acción muscular.
En la porción posterior de la superficie abdominal de la araña hay un grupo de glándulas "hileras", apéndices modificados de los cuales se exuda una proteína fluida que se polimeriza formando seda cuando se expone al aire. La seda se usa, no sólo para tejer las telas con las que atrapan a las presas, sino para otros propósitos como para hacer una línea de descenso por la cual la araña puede hacer un escape defensivo, una cuerda guía para indicar un curso, un hilo finísimo para aterrizar, bisagras para puertas trampa, sacos de huevos, un tapiz para sus nidos o un recubrimiento de sus presas o del alimento que los machos de ciertas especies ofrecen al cortejar a las hembras.

Una araña.
Los conductos de las glándulas de veneno se abren en los extremos de
los quelíceros o próximos a éstos. El flujo de veneno es
controlado voluntariamente por la araña. Sólo unas pocas arañas
son peligrosas para los seres humanos; tal vez las más peligrosas son
los miembros de la especie conocida comúnmente como "viuda negra".
Las 25.000 especies de crustáceos incluyen cangrejos, langostinos, langostas, camarones, centollas, percebes, pulgas de agua (Daphnia) y algunas formas más pequeñas. Algunos crustáceos, tales como los conocidos bichos bolita o cochinillas de tierra, están adaptados a la vida en ambientes terrestres húmedos. Los crustáceos difieren de los mandibulados terrestres -como los insectos que veremos más adelante- en que tienen patas, o apéndices semejantes a patas, en el abdomen al igual que en el tórax, y tienen dos pares de antenas en comparación con el único par de los insectos. Muchos presentan caparazones. En los crustáceos, los sexos habitualmente están separados.
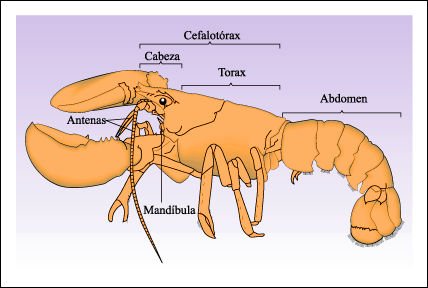
Un crustáceo representativo, el bogavante americano, Homarus americanus.
El bogavante tiene dos pares de antenas (que no son verdaderos apéndices)
y 17 pares de apéndices que incluyen piezas bucales y patas especializadas
en la alimentación, locomoción y natación. Los primeros
13 segmentos se unen dorsalmente en una combinación de cabeza y tórax
-el cefalotórax-. Un pesado escudo o caparazón surge desde la
cabeza y cubre el tórax. El abdomen consiste en 6 segmentos -diferenciados
pero similares- y un telson terminal, que tiene el ano en su base. Los apéndices
tienen distintas funciones: las antenas tienen funciones sensoriales; las mandíbulas,
la maxilula y la maxila trituran el alimento. Como en todos los artrópodos,
las mandíbulas se mueven lateralmente, abriéndose y cerrándose
de lado a lado como un par de pinzas de hielo. El cefalotórax tiene 3
pares de maxilipedios y 5 pares de patas locomotoras o pereiopodos, de las cuales
el primer par está modificado y forma pinzas (una de mayor tamaño
para defensa y para triturar alimento y otra más pequeña, con
dientes más filosos, que atrapa y desmenuza la presa). Los otros 2 pares
de patas locomotoras tienen pequeñas pinzas que también pueden
capturar presas. El último par de patas locomotoras, los pleópodos,
sirven también para limpiar los apéndices del abdomen y como aletas
en la natación. Las pinzas y el abdomen presentan músculos estriados
grandes muy poderosos. Cualquiera de los apéndices puede regenerarse
al cabo de una serie de mudas si se pierde y, frecuentemente, el bogavante es
capaz de seccionar (autotomizar) un apéndice atrapado por un depredador
para poder escapar.
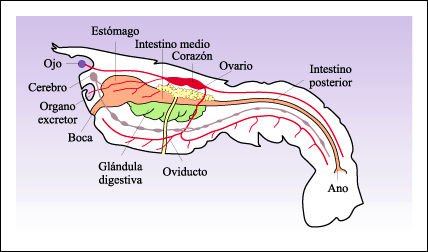
Diagrama que representa la anatomía interna del bogavante.
En el tubo digestivo, el intestino
anterior y el posterior están revestidos de cutícula. En consecuencia,
la mayor parte del alimento se absorbe a través del intestino medio y
de las células del hepatopáncreas, una gran glándula digestiva
que se origina a partir de un par de divertículos del intestino medio.
La respiración se realiza cuando fluye el agua a través de los
20 pares de branquias plumosas, insertas en o cerca de la base de las patas.
En la cabeza hay un par de órganos excretores; los desechos extraídos
de la sangre son colectados en una vejiga y excretados por un poro en la base
de cada una de las segundas antenas.
A diferencia de otros grupos
de artrópodos, casi todos los crustáceos son acuáticos;
sin embargo, algunos cangrejos son anfibios o terrestres. Los cangrejos anfibios
continúan respirando por branquias, llevando agua en sus cavidades torácicas
con la que mantienen las branquias húmedas. Los cangrejos terrestres
verdaderos carecen de algunas de las estructuras branquiales, pero poseen un
área de tejido epitelial muy vascularizado a través del cual se
intercambian los gases.
Los mandibulados terrestres
se identifican por su único par de antenas y por sus mandíbulas
las que difieren de las de los crustáceos. Respiran a través de
tráqueas y su excreción se efectúa por medio de túbulos
de Malpighi. Además de los insectos, hay cuatro clases más pequeñas
de mandibulados terrestres, relativamente poco especializados. En estos artrópodos
ha habido poca tagmosis y sus cuerpos consisten en una región cefálica,
seguida de un tronco alargado con muchos segmentos diferenciados, todos más
o menos semejantes. Con pocas excepciones, todos los segmentos tienen apéndices
pares. Los miembros de estas cuatro clases se conocen colectivamente como miriápodos
("con muchos pies").
Los miriápodos más
familiares son los ciempiés, clase Chilopoda (Quilópodos) y los
milpiés, clase Diplopoda (Diplópodos). Las aproximadamente 3.000
especies de ciempiés tienen preferencia por los lugares húmedos,
debajo de los troncos o rocas, o en sótanos. Todos son carnívoros,
se alimentan de cucarachas y otros insectos, así como de anélidos
de cuerpo blando.
Los insectos son los únicos
invertebrados capaces de volar.
Hay aproximadamente 30 órdenes de insectos, de los cuales los cuatro mayores son Diptera, Lepidoptera, Hymenoptera y Coleoptera. Los dípteros ("con dos alas") incluyen las moscas, jejenes y mosquitos. Los lepidópteros ("con alas escamosas") son las polillas y las mariposas. Los himenópteros ("con alas membranosas") incluyen hormigas, avispas y abejas, muchas de cuyas especies viven en sociedades complejas. La mayoría posee un par de alas anteriores protectoras duras, que permanecen elevadas durante el vuelo, y un par de alas posteriores membranosas que usan para impulsarse.
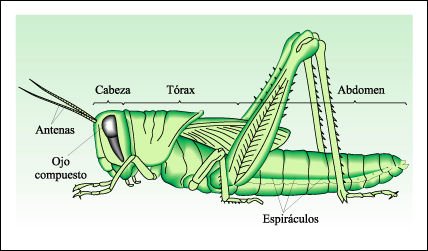
Un saltamontes.
En el saltamontes, un insecto, la cabeza está formada por seis segmentos
fusionados que tienen apéndices especializados en la gustación
y la masticación. Cada uno de los tres segmentos del tórax tiene
un par de patas (tres en total) y los dos primeros llevan un par de alas cada
uno(en el saltamontes las alas anteriores están semiendurecidas como
cubiertas protectoras de las alas posteriores membranosas). Los espiráculos
del abdomen se abren en una red de túbulos revestidos de cutícula,
a través de los cuales circula el aire a los distintos tejidos del cuerpo.
Este tipo de sistema respiratorio tubular se encuentra en los mandibulados terrestres
y en algunos arácnidos. La excreción se realiza a través
de los túbulos de Malpighi que se vacían en el intestino posterior.

Piezas bucales de un saltamontes.
Las mandíbulas son apéndices trituradores. El labium y el labrum
son los labios inferior y superior. Las maxilas hacen pasar el alimento a la
boca y los palpos contribuyen a la gustación y manejo del alimento.

Fotografía de un saltamontes comiendo. Nótese cómo utiliza los palpos en el manejo del alimento.
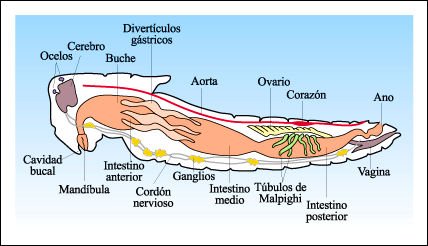
La anatomía interna de un insecto, ejemplificada en el saltamontes.
Los ciclos biológicos
de los insectos son fundamentalmente diferentes de aquellos de los invertebrados
marinos, particularmente de las formas que cuando alcanzan el estado adulto
son sésiles o de movimiento lento. Aunque muchas larvas de invertebrados
marinos se alimentan, su principal actividad parece ser la invasión y
selección de nuevos hábitat a cierta distancia del hábitat
del progenitor. Estas larvas pueden migrar a grandes distancias y esto resulta
en una dispersión de la especie en un área amplia.
Las formas inmaduras de la mayoría
de los insectos, en contraste, tienen una movilidad limitada y, habitualmente,
pasan a través de las distintas etapas del desarrollo muy cerca del lugar
donde la hembra depositó originalmente los huevos, a veces exactamente
en ese mismo lugar. La capacidad de volar les ha conferido a la mayoría
de los insectos adultos una movilidad extraordinaria. Una ventaja evidente del
vuelo es el hecho de facilitar la localización de una pareja o de nuevos
hábitat en los cuales depositar sus huevos, asegurando así la
dispersión.
En todos los insectos los sexos
están separados. En muchos de ellos se ha desarrollado un cierto grado
de cuidado de la cría por parte de los progenitores por lo que uno de
ellos, o ambos, protegen o alimentan a la progenie, o hacen ambas tareas a la
vez. Entre las termitas, hormigas, avispas y abejas, este cuidado se encuentra
en un contexto social complejo, en el que es característica una división
sustancial del trabajo.
Los insectos jóvenes
en crecimiento a menudo sufren una metamorfosis. El grado de cambio es diferente
en los distintos órdenes de insectos. En algunas especies, los jóvenes,
aunque sexualmente inmaduros, se parecen a adultos pequeños y aumentan
de tamaño por una serie de mudas, hasta que alcanzan su tamaño
máximo. En el saltamontes, por ejemplo, los recién nacidos son
ápteros -sin alas- y las alas comienzan a desarrollarse en etapas inmaduras
posteriores; alcanzando su total desarrollo en el estadio adulto, en el que
son funcionales. En todos los otros aspectos son similares al adulto. Estas
formas inmaduras, incapaces de reproducirse, se conocen como larvas. Se dice
que los insectos que alcanzan el estadio adulto gradualmente con las sucesivas
mudas tienen metamorfosis incompleta. Sin embargo, casi el 90% de las especies
de insectos presentan una metamorfosis completa y los adultos son drásticamente
diferentes de las formas inmaduras. Estas formas de alimentación inmaduras
-las larvas- se conocen comúnmente como orugas. Después del período
larval, el insecto sufre la metamorfosis completa y pasa a un estado de pupa,
en el cual ocurre una remodelación extensa del organismo. El insecto
adulto (maduro sexualmente) emerge de la pupa. En muchos casos, los huevos o
las pupas (que no se alimentan) son capaces de soportar épocas desfavorables,
como por ejemplo, estaciones frías o secas.

Ciclo biológico de Heliconius ismenius , mariposa de América Central. Su ciclo vital comienza con el apareamiento de mariposas macho y hembra.

La fecundación es interna y los óvulos fecundados pasan las etapas tempranas de su desarrollo dentro de una vaina ovígera gomosa.

La oruga recién nacida, o forma larval, come, crece y continúa su desarrollo.

Tercera intermuda, o sea, la oruga después de su segunda ecdisis.

Cuando está lista para pasar a pupa, la oruga se suspende boca abajo de una hoja mediante un penacho de seda.

Dentro de la pupa ocurre la metamorfosis a mariposa, a través de la remodelación de los tejidos de la oruga. La mariposa adulta que emerge es igual a a), es la forma de reproducción sexual, lista para comenzar el ciclo una vez más.
La primera forma es el embrión dentro del huevo. La segunda forma es
la larva, el animal que nace del huevo; las larvas se alimentan y crecen. Durante
el curso de su crecimiento, la larva muda un número característico
de veces. Luego, cuando la larva ha crecido completamente, muda y forma la pupa.
En muchas especies, pero no en todas, la pupa está encerrada en un capullo
u otra cubierta protectora. Durante la etapa de pupa, que externamente parece
inerte, muchas de las células larvales se desintegran y grupos de células
completamente nuevas, que se encontraban en reserva en el embrión, comienzan
a proliferar y usan el tejido larval en degeneración como medio de cultivo.
Estos grupos de células desarrollan las estructuras complejas características
del adulto, tales como las alas o los ojos compuestos, entre otras.
La muda y la metamorfosis están
bajo el control hormonal.Al igual que la mayoría de los procesos controlados
por hormonas, la muda y la metamorfosis son el resultado final de la interacción
de varias sustancias: la hormona cerebral, la hormona de la muda -o ecdisona-
y la hormona juvenil.
Razones del éxito de
los artrópodos
¿Por qué los artrópodos,
en general, y los insectos, en particular, han tenido un éxito tan espectacular
entre los invertebrados? Una razón importante es, sin duda, la naturaleza
del exoesqueleto, que es impermeable al agua, provee protección y hace
posible la evolución de los muchos apéndices finamente articulados,
característicos de este phylum.
Otra razón, que se aplica
especialmente a los insectos, es su pequeño tamaño, la alta especificidad
de la dieta y otros requerimientos de cada especie. En consecuencia, muchas
especies diferentes pueden vivir en un único ambiente pequeño
sin competir unas con otras. Las piezas bucales variadas y altamente especializadas
son un reflejo de esta especificidad en la dieta.
La diversidad de los insectos
y la especificidad de sus requerimientos puede ser, en parte, una respuesta
evolutiva a la gran diversidad de microambientes proporcionados por las plantas
vasculares. Hay sólo 235.000 especies de angiospermas pero, además,
la estructura de cada planta es tan compleja que suministra una variedad de
recursos que pueden ser explotados por insectos con diferentes requisitos y
adaptaciones. Éste es un ambiente mucho más rico que el de altamar,
las costas marinas o el suelo. Los recursos ofrecidos por las plantas vasculares
fueron, en efecto, un laboratorio evolutivo en el que pudieron ensayarse numerosísimas
variaciones, que originaron la diversidad de adaptaciones que vemos actualmente.
Este proceso, por supuesto, continúa.
Otro factor del éxito
de los insectos es la metamorfosis completa que ocurre en la vasta mayoría
de sus especies. En ellas, las adaptaciones para la alimentación y el
crecimiento que se encuentran en las larvas, están separadas de las adaptaciones
para la dispersión y la reproducción que se encuentran en los
adultos. Otra consecuencia de la metamorfosis completa es que los requerimientos
dietarios y otros de las larvas y de los adultos de la misma especie son tan
diferentes que no compiten entre sí. En efecto, ocupan ambientes diferentes.
Una razón final del éxito
es, indudablemente, el sistema nervioso de los artrópodos con su delicado
control sobre los varios apéndices y los muchos órganos sensoriales
altamente sensibles que se encuentran en gran diversidad a través del
phylum.
El órgano sensorial más conspicuo de los artrópodos es el ojo compuesto, que es característico de este phylum. Los ojos compuestos están formados por múltiples estructuras de enfoque, cada una de ellas asociada a un número reducido de células fotorreceptoras; cada unidad constituye un ommatidio. El ommatidio es la unidad estructural básica del ojo compuesto.
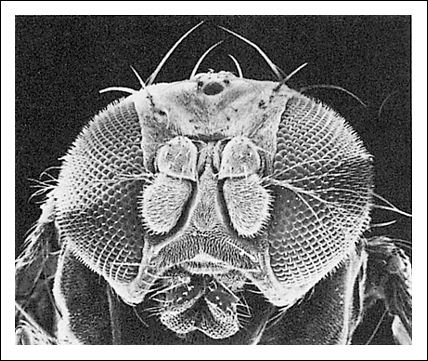
La cabeza de Drosophila, como se ve con el microscopio electrónico de barrido.
Nótense los grandes ojos compuestos a cada lado de la cabeza que se observan
en la fotografía. Aunque los ojos del insecto no pueden cambiar de foco,
pueden definir objetos situados a sólo un milímetro de distancia
de la lente, adaptación útil para un insecto.
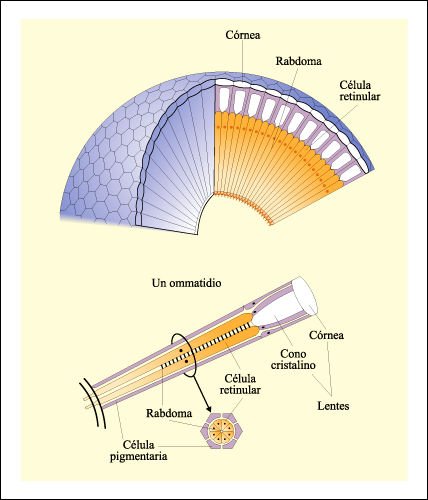
Estructura del ojo compuesto.
El ojo compuesto está formado por un gran número de unidades estructurales
y funcionales llamadas ommatidios. Cada ommatidio tiene su propia lente, la
córnea, que forma una de las facetas del ojo compuesto, además
de otra lente, el cono cristalino. Ambas lentes enfocan la luz. La parte de
las células retinulares sensible a la luz es el rabdoma. El estímulo
es transmitido por los axones de las células retinulares al sistema nervioso
central. El ommatidio está rodeado por células pigmentarias que
impiden que la luz viaje de un ommatidio a otro.
Los artrópodos tienen
además otros dos tipos de órganos visuales con distinto origen
embrionario: los ocelos dorsales -exclusivos de algunos insectos adultos y coexistentes
con los ojos compuestos- y los ojos simples. Los ojos simples, "tipo cámara"
se caracterizan por poseer una única estructura de enfoque -o lente-
compartida por la totalidad de los fotorreceptores que forman la retina. Desde
el punto de vista funcional, los ocelos dorsales se caracterizan por estar desenfocados.
Por esta razón, no forman imagen, algo que sí ocurre en los ojos
simples. Esta incapacidad de los ocelos permite, sin embargo, diferenciar, por
ejemplo, el área por encima del horizonte (cielo) de la que se encuentra
por debajo (tierra). De esta manera, funcionan como detectores del horizonte
y, de este modo, pueden asistir a la estabilización del vuelo.
La superficie corporal de los
artrópodos terrestres frecuentemente está cubierta por unidades
receptoras sensoriales conocidas como sensilias -"pequeños órganos
sensoriales"-. La mayoría de las sensilias tienen forma de espinas
finas o sedas, compuestas por columnas huecas de cutícula.
Los propioceptores son receptores sensoriales que proveen información acerca de la posición de distintas partes del cuerpo y de las tensiones y presiones que se ejercen sobre ellas. Un tipo de sensilia común en los artrópodos es la sensilia campaniforme.
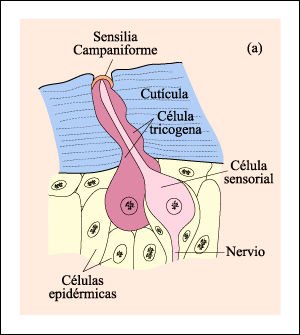
Sensilias campaniformes, un tipo de receptores de estímulos mecánicos comunes en los artrópodos.

Mecanismo de la Mantis religiosa para localizar a un insecto
Dado que sus ojos
no se mueven, la Mantis religiosa debe mover su cabeza entera para tener a su
víctima en visión binocular. El movimiento de su cabeza envía
impulsos a las patas a través de pelos propioceptivos de la cabeza y
el tórax. El movimiento de sus patas es así automáticamente
coordinado con la posición de su presa, confiriendo a la mantis la capacidad
de apresar velozmente a una víctima con un zarpazo adecuado.Los artrópodos,
particularmente los insectos, presentan formas complejas de comunicación.
Varias especies, tales como las langostas, los saltamontes y los grillos, se
llaman unos a otros con sonidos producidos al raspar entre sí sus patas
o sus alas, o al frotarlas contra sus cuerpos. El reconocimiento de un sonido
puede basarse en su patrón, su ritmo o su frecuencia. Los insectos producen
cantos y responden a melodías apropiadas sin haberlas escuchado previamente.
Los receptores
de sonido más simples en un artrópodo son las sensilias con pelos
táctiles que vibran cuando las golpean las moléculas del aire
en movimiento; a su vez, las vibraciones de estos pelos disparan impulsos eléctricos
en el sistema nervioso del animal.
Otros insectos tienen estructuras especiales que responden a los cambios de presión en las ondas sonoras; estas estructuras, que pueden estar localizadas sobre las patas, el tórax o el abdomen, se conocen como órganos timpánicos. En estos órganos, una membrana fina, el tímpano, se extiende a través de uno o más sacos aéreos cerrados.
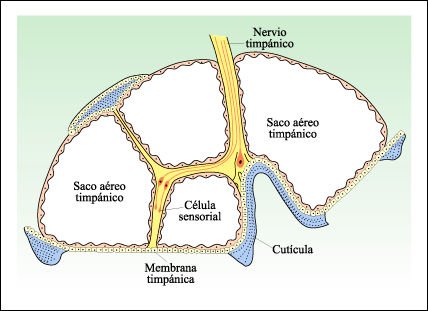
El receptor sonoro más complejo de los insectos es el órgano timpánico.
Los sacos aéreos timpánicos están cubiertos por un tambor
membranoso y las células sensoriales están dispuestas de tal manera
en el órgano, que son estimuladas por movimientos del tambor o las paredes
del saco aéreo. Los órganos timpánicos responden a cambios
de presión del medio. Esto es un corte transversal del órgano
timpánico de una polilla noctuida.
Algunas de las muchas sensilias
que podemos encontrar en un insecto presentan pequeños poros sobre su
superficie. A través de estos poros, moléculas presentes en el
entorno se pueden unir a sitios específicos de reconocimiento, ubicados
en la membrana de las células asociadas a tales sensilias. Los receptores
químicos pueden cumplir funciones de detección de sustancias presentes
sobre una superficie -quimiorreceptores de contacto- o disueltos en el aire
o el agua -quimiorreceptores olfativos-. Los primeros pueden estar asociados
al reconocimiento del alimento -por lo que a veces son llamados "gustativos"-
o a otras señales, tales como marcas que dejan algunos insectos sobre
ciertas superficies o sendas. Los quimiorreceptores olfativos a veces están
especializados en la detección de compuestos variados, incluyendo alimento
y señales de comunicación, y se localizan mayormente en las antenas
del insecto.
Muchos insectos se comunican
por sustancias químicas conocidas como feromonas.


