
Capítulo 34.
El reino
animal IV: los deuteróstomos
Cuatro grupos de animales tienen desarrollo deuteróstomo; ellos son:
Echinodermata, Chaetognatha, Hemichordata y Chordata.
Los equinodermos incluyen los
lirios de mar y las estrellas pluma, las estrellas de mar, las estrellas frágiles,
los erizos de mar y los dólares de arena, y los pepinos de mar. Aunque
los equinodermos tienen larvas con simetría bilateral, las formas adultas
de la mayoría de las especies tienen simetría radial, con un plan
corporal pentámero.
El phylum Chaetognatha comprende
a los gusanos flecha. Los gusanos flecha tienen tres regiones corporales distintas
y están entre los predadores más activos del plancton marino.
Los hemicordados comprenden
convencionalmente a los pequeños Pterobranchia -formas sésiles
con tentáculos- y a los Enteropneusta -de vida libre con forma de gusano-.
Sin embargo, la relación entre ambos grupos no es completamente clara.
Los hemicordados presentan características que comparten con equinodermos
y cordados. Sus larvas, las cavidades celómicas de algunos miembros del
phylum, y porciones del sistema nervioso se asemejan a las de los equinodermos.
Sin embargo, los hemicordados comparten el cordón nervioso dorsal (no
ventral) con los cordados, así como una faringe con hendiduras branquiales.
El phylum Chordata comprende
tres subphyla: Cephalochordata (peces lanceta), Urochordata (tunicados) y Vertebrata
(vertebrados). Las características fundamentales de los cordados son
la notocorda -una varilla longitudinal flexible que corre ventralmente con respecto
al cordón nervioso y sirve como eje estructural del cuerpo (presente
sólo en la vida embrionaria de la mayoría de los vertebrados)-,
el cordón nervioso -un tubo hueco localizado dorsalmente- y la presencia
de cola.
Los vertebrados, el mayor grupo
de cordados, se caracterizan por una serie de rasgos en la región cefálica;
en la gran mayoría, se desarrolla una columna vertebral y un cráneo
que protege al cerebro. Los vertebrados incluyen a los peces (tres clases vivientes),
los anfibios, los reptiles, las aves y los mamíferos.
Algunos peces óseos devónicos,
precursores de los anfibios, se adaptaron a vivir en aguas poco profundas y
esos caracteres adaptativos -incluyendo la presencia de pulmones y el reforzamiento
de las estructuras esqueletarias- parecen haber favorecido su transición
a la tierra. La mayoría de los anfibios modernos están adaptados
en forma incompleta a la vida terrestre y deben pasar parte de su ciclo vital
en el agua. La aparición del huevo amniota en un grupo de tetrápodos
posibilitó que los vertebrados conquistaran más completamente
los ambientes terrestres. La diversificación de los amniotas finalmente
originó no sólo una gran variedad de reptiles (muchos de los cuales
se extinguieron hace aproximadamente 65 millones de años) y sinápsidos
no mamíferos, sino también sus descendientes: las aves y los mamíferos.
Phylum Echinodermata (equinodermos):
animales de "piel espinosa"
Los equinodermos incluyen las
estrellas pluma y los lirios de mar (crinoideos), las estrellas de mar (asteroideos),
las estrellas frágiles (ophiuroideos), los erizos de mar y los dólares
de arena (echinoideos) y los pepinos de mar (holothuroideos).
La mayoría de los equinodermos
adultos tienen simetría radial, pero con algunos indicios de bilateralidad.
Sin embargo, las larvas presentan simetría bilateral. Durante el desarrollo,
diferentes partes del cuerpo de la larva crecen a diferentes velocidades, preparando
el camino para los cambios en la simetría del cuerpo.
En el momento de la metamorfosis,
las larvas se fijan temporariamente a una superficie sólida; luego se
transforman rápidamente en adultos con una simetría radial pentámera.
Se cree que los equinodermos han evolucionado a partir de una forma móvil
de simetría bilateral, que posteriormente se adaptó a una vida
sésil. Ese estadio fijo en la evolución de los equinodermos, que
tendría alimentación filtrante, se habría beneficiado con
la adquisición de una simetría radial. Esta simetría se
habría desarrollado alrededor del punto de fijación y habría
dejado al organismo igualmente expuestoa
los factores del medio.
Subsecuentemente, algunos equinodermos
- como las estrellas de mar y los erizos de mar- podrían haberse vuelto
móviles nuevamente. Siguiendo esta línea de razonamiento, uno
podría esperar un probable regreso a la simetría bilateral en
este grupo y, efectivamente, esto se ve hasta cierto punto en los cuerpos alargados,
blandos de los pepinos de mar.
Los rasgos característicos de los equinodermos se ven claramente en uno de los representantes más familiares: la estrella de mar, aunque es posible que ese plan de organización se encuentre modificado con respecto al de los primitivos equinodermos.
El plan corporal de cinco partes, bien visible en la estrella de mar que se muestra aquí, es característico de la mayoría de los equinodermos vivientes.
Los equinodermos se caracterizan por un esqueleto calcáreo interno, de
origen mesodérmico, formado por placas que toman la forma de una red
tridimensional, un sistema vascular acuífero de canales y pies ambulacrales.
El sistema vascular acuífero es una característica única de todos los equinodermos. Este sistema consiste en una red de tubos cuya cavidad de origen celómico es hidrostáticamente continua con el agua circundante. Estos tubos conducen a los pies ambulacrales, peculiares estructuras musculares que cumplen diferentes funciones. En las estrellas de mar, cumplen una función locomotora

La locomoción en las estrellas de mar es llevada a cabo por el sistema vascular acuífero.
a) Cinco conductos radiales, uno para cada brazo, conectan el conducto anular
con numerosos pares de pies ambulacrales -cilindros huecos, de paredes gruesas,
que terminan en ventosas-.Las ventosas están en el extremo de cada pie
ambulacral y por medio de ellas se adhieren al sustrato. b) En el otro extremo
se encuentra un saco muscular redondeado, la ampolla. c) Cuando la ampolla se
contrae, el agua que contiene -que no puede volver al canal radial porque una
válvula se lo impide- entra a presión en el pie ambulacral. Esto
endurece y estira al pie, otorgándole suficiente rigidez como para sostener
la marcha. d) Los músculos del pie se contraen después, obligando
al agua a retornar a la ampolla y originar la succión que mantiene el
pie adherido al sustrato. Cuando los pies ambulacrales se implantan sobre una
superficie dura, tal como una roca o la concha de una almeja, las contracciones
de los músculos en la base de cada pie ejercen suficiente fuerza colectivamente
como para que la estrella de mar avance o pueda abrir las valvas de un molusco
bivalvo. Cuando ataca a los bivalvos, que son su dieta básica, la estrella
de mar evierte su estómago a través de su abertura bucal e introduce
el tejido estomacal en la abertura que ha logrado formar entre las conchas del
bivalvo. Los tejidos estomacales pueden insinuarse a través de una hendidura
de hasta 0,1 milímetro para digerir el tejido blando de la presa.
Cada brazo contiene un par de
glándulas digestivas y también un cordón nervioso, con
una mancha ocular en su extremo. Estas manchas oculares son los únicos
órganos sensoriales, estrictamente hablando, de la estrella de mar, pero
la epidermis contiene miles de células neurosensoriales vinculadas al
tacto, la fotorrecepción y la quimiorrecepción. Los sexos están
separados en la mayoría de los equinodermos, pero algunos asteroideos
son ocasionalmente hermafroditas.
La respiración se lleva
a cabo por numerosas proyecciones dactiliformes pequeñas -las branquias
dérmicas- protegidas por espinas. La eliminación de los desechos
está a cargo de células ameboides que circulan en el fluido celómico
capturando los desperdicios y escapando luego a través de las delgadas
paredes de las branquias dérmicas, donde esos despedicios son eyectados.
Phylum Chaetognatha (quetognatos):
gusanos flecha
Los gusanos flecha no son segmentados,
pero sus cuerpos, de simetría bilateral, tienen tres regiones distintas:
cabeza, tronco y cola; las dos últimas regiones presentan aletas de origen
epidérmico reforzadas por radios. Su desarrollo embrionario los identifica
como deuteróstomos, pero no parecen estar cercanamente relacionados con
ningún otro phylum de deuteróstomos.
Todos los gusanos flecha son
hermafroditas y, en algunas especies, ocurre autofecundación. La progenie
joven se asemeja a adultos en miniatura y, sin pasar por un estado larval diferente,
comienzan con rapidez una vida de depredación activa.
Los gusanos flecha son un componente importante del plancton marino; se alimentan de copépodos y, en ocasiones, de pequeños peces. Habitualmente se alimentan cerca de la superficie durante la noche, descendiendo a aguas más profundas durante el día.
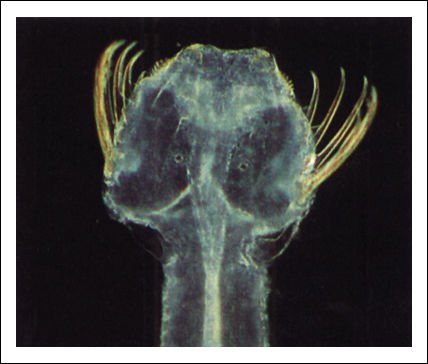
Un gusano flecha está maravillosamente equipado para sus actividades depredadoras.
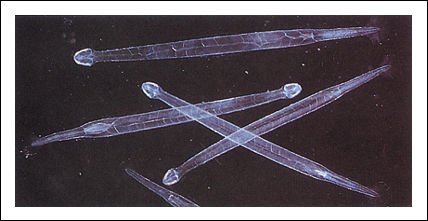
Gusanos flecha.
La cabeza tiene dos ojos grandes en la superficie dorsal y numerosas espinas
agudas, que usa para arponear a la presa. Dentro de la cámara que lleva
a la boca se encuentran fuertes dientes.
Phylum Hemichordata (hemicordados):
gusanos bellota
Los hemicordados tienen particular
interés desde el punto de vista evolutivo porque tienen rasgos característicos
de los equinodermos y de los cordados. Algunos de los hemicordados tienen larvas
ciliadas que son casi idénticas a las larvas de la estrella de mar. Además,
las cavidades celómicas de los tentáculos de los pterobranquios
suministran un soporte hidrostático semejante al sistema vascular acuífero
de los equinodermos, aunque no presentan pies ambulacrales.
El sistema nervioso de los hemicordados
es, en algunos aspectos, similar al de los equinodermos, pero incluye cordones
nerviosos ventral y dorsal que están unidos por un anillo en el límite
posterior del collar. En el resto del reino animal, sólo en los cordados
existe un cordón nervioso principal en el lado dorsal del cuerpo. El
cordón nervioso dorsal de los cordados es hueco, a diferencia de los
cordones nerviosos ventrales de otros animales, que son sólidos. En algunos
hemicordados, la porción anterior del cordón dorsal también
es hueca.
La evidencia más fuerte
de una relación estrecha entre los hemicordados y los cordados la proporciona
la faringe, una estructura de la porción anterior del tronco, que está
perforada por orificios denominados hendiduras branquiales. El tejido muy vascularizado
que circunda a las hendiduras sirve como aparato de intercambio de gases respiratorios,
mientras que los orificios permiten la salida del agua que entra por la boca.
Como veremos, esta faringe perforada también está presente en
los cordados.
Todos los hemicordados son marinos. Los Pterobranchia forman parte del bentos de aguas profundas; los Enteropneusta son cavadores en aguas más superficiales. En los enteropneustos el cuerpo está dividido en tres regiones: una probóscide, con la cual el animal excava en los sedimentos oceánicos, un collar corto y un tronco largo. Los pterobranquios tienen las mismas tres regiones corporales y comparten otras características con los enteropneustos.

Un gusano bellota, Glossobalanus sarniensis enterrándose en una grava de conchas.
El cuerpo del gusano bellota,
Glossobalanus sarniensis sarniensis consiste en tres regiones: una probóscide,
un collar corto y un tronco largo, sólo una porción del cual es
visible aquí. Nótense las hendiduras branquiales en la porción
anterior del tronco. Estas hendiduras se hallan también presentes en
los cordados.
Phylum Chordata (cordados):
los cefalocordados y los urocordados
El phylum Chordata incluye aproximadamente
43.000 especies agrupadas en tres subgrupos: los Cephalochordata -también
llamados lancetas o anfioxos-, los Urochordata -o tunicados- los más
comunes de los cuales son las papas de mar, y los Vertebrata -o vertebrados-.
Así, podemos ver que el vocablo "invertebrado" se refiere a
todos los animales, con excepción de los miembros de un subgrupo de cordados.
Cephalochordata incluye sólo
alrededor de 28 especies. El cefalocordado mejor conocido es Branchiostoma,
el anfioxo.
Otro grupo de cordados es el
de los Urochordata. Aunque los urocordados adultos no tienen todos los rasgos
típicos de los cordados, las larvas -que se asemejan a Branchiostoma
y tienen vida libre durante un tiempo muy corto- sí poseen esas características.
Por esta razón, se ha propuesto que el ancestro común de cefalocordados
y vertebrados pudo haberse originado a partir de larvas de urocordados que alcanzaron
la madurez sexual reteniendo los rasgos larvales. Este mecanismo se denomina
paedomorfosis. Se conocen aproximadamente 1.300 especies de urocordados, que
se encuentran en el plancton y en el fondo de los océanos de todo el
mundo, tanto en aguas de escasa profundidad como profundas. Se conocen comúnmente
como tunicados, porque el cuerpo está cubierto por una túnica
protectora y firme que contiene una sustancia similar a la celulosa.
El anfioxo muestra las características distintivas de los cordados 1) una notocorda, varilla dorsal que se extiende a lo largo del cuerpo; 2) un cordón nervioso dorsal tubular y 3) una cola. Además tienen hendiduras branquiales que también presentan los hemicordados. Branchiostoma retiene estas características durante toda su vida; sin embargo, muchos cordados tienen una notocorda, hendiduras o bolsas branquiales faríngeas y una cola sólo durante sus etapas inmaduras.
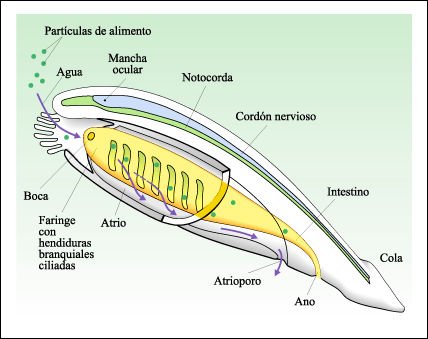
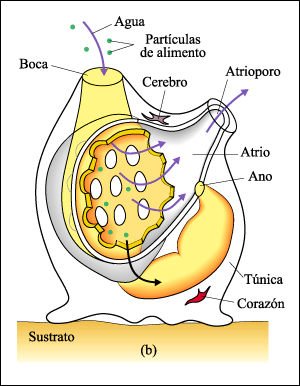
El anfioxo Branchiostoma

Peces lanceta con su parte anterior emergiendo del sustrato.
Gran parte del tejido corporal está formado por bloques de músculos
segmentarios. La hilera de estructuras cuadradas, amarillentas a lo largo del
costado del cuerpo son los órganos reproductores. En los peces lanceta
los sexos están separados, y los espermatozoides o los óvulos
se vierten en el atrio. Los gametos salen a través del atrioporo, y la
fecundación ocurre externamente.
Los tunicados tienen dos etapas en su ciclo de vida: la larva y el adulto.
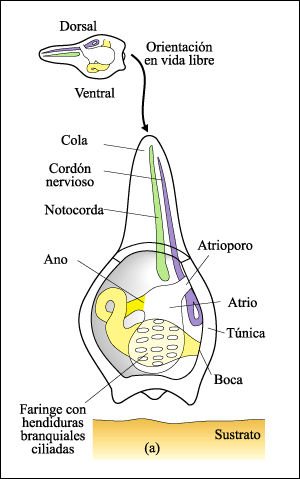
a) Larva de un tunicado.
En la larva, la túnica cubre la boca y el atrioporo, lo que evita el
flujo de agua a través de la faringe y de sus hendiduras branquiales
ciliadas. Así, inclusive en las especies en las cuales la faringe larval
está bien desarrollada, como se muestra aquí, la larva es incapaz
de alimentarse.
b) Forma adulta de un tunicado.
Después de una breve existencia libre, se asienta en el fondo y se fija
por el extremo anterior. La metamorfosis comienza entonces. Desaparece la cola
de la larva con la notocorda y el cordón nervioso dorsal, y el cuerpo
entero del animal presenta un giro de 180 grados. La boca es llevada hacia atrás
y se abre en el extremo opuesto al de fijación, y todos los otros órganos
internos también rotan hacia atrás. Se ha supuesto que los vertebrados
ancestrales surgieron de larvas tunicadas que se hicieron sexualmente maduras
y, así, fueron capaces de reproducirse, sin sufrir metamorfosis.
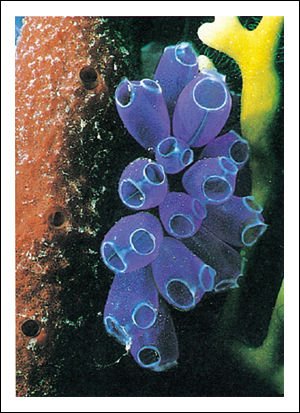
Tunicados adultos vivos: papas de mar o ascidias
Phylum Chordata (cordados): los vertebrados
Los vertebrados constituyen
el subgrupo más grande y más conocido de cordados. Ciertas características
que presentan en la región cefálica, como la concentración
de órganos sensoriales y el cerebro, distinguen claramente a los vertebrados,
aun a los más primitivos, de los cordados no vertebrados.Se
ha sugerido que el pasaje de un modo pasivo de alimentación -por filtración
ciliar- a otro activo fue acompañado de cambios en la actividad muscular
y cefalización que diferencian a los ancestros de los vertebrados de
los vertebrados actuales.
La mayoría de los vertebrados
tiene una columna vertebral -o espina dorsal- como eje estructural; sin embargo
las formas más primitivas carecen de vértebras. La columna vertebral
es un soporte flexible, habitualmente óseo, que se desarrolla alrededor
de la notocorda, suplantándola por completo en la mayoría de las
especies. Las proyecciones dorsales de las vértebras rodean al cordón
nervioso a lo largo de la columna. El cerebro está, de modo semejante,
encerrado y protegido por un cráneo. Entre las vértebras hay discos
cartilaginosos que dan flexibilidad a la columna vertebral. En asociación
con las vértebras, hay músculos segmentados por medio de los cuales
sectores de la columna pueden moverse por separado. Este patrón segmentado
persiste en las formas embrionarias de los vertebrados superiores, pero se pierde
por completo en el curso del desarrollo.
Una de las grandes ventajas
de un endoesqueleto óseo es que está formado por tejido vivo que
puede crecer con el animal. En el embrión vertebrado en desarrollo, el
esqueleto es principalmente cartilaginoso; en la mayoría de los vertebrados,
los huesos reemplazan gradualmente al cartílago en el curso de la maduración.
Las porciones en crecimiento de los huesos siguen siendo, característicamente,
cartilaginosas, hasta que el animal alcanza su tamaño definitivo.
Hay siete grupos vivientes de
vertebrados: los peces agnatos, óseos y cartilaginosos, los anfibios,
los reptiles, las aves y los mamíferos. Su evolución está
claramente documentada en el registro fósil.
Los primeros vertebrados carecían
de mandíbulas y la notocorda no era reemplazada por la columna vertebral.
Actualmente, sólo los mixines y las lampreas representan este estadio
de la evolución de los vertebrados. Sin embargo, los peces sin mandíbula
fueron muy diversos, como lo muestra su registro fósil.
Uno de los principales acontecimientos
en la evolución de los vertebrados fue la aparición de las mandíbulas.
Estas estructuras se originaron como producto de modificaciones del patrón
de desarrollo de la región cefálica anterior y constituyeron una
verdadera novedad evolutiva. El desarrollo de mandíbulas, frecuentemente
dotadas de dientes, amplió la variedad de organismos que les servían
de alimento. Junto con una alimentación más eficiente obtenida
de fuentes de energía más grandes y más concentradas, en
los gnatostomados (vertebrados con mandíbulas) se abrió la posibilidad
de incrementos significativos de tamaño.
Los tiburones y las rayas se incluyen entre los condrictios, la segunda clase importante de peces. Presentan un esqueleto cartilaginoso, aunque es posible que sus antecesores, como los de los agnatos vivientes, fueran animales capaces de formar hueso. La piel de los condrictios está cubierta por escamas denticuliformes pequeñas y puntiagudas (escamas placoides) que no se superponen y se asemejan estructuralmente a los dientes de los vertebrados. Estas escamas le dan a la piel la textura y la calidad abrasiva de un papel de lija grueso. Algunos placodermos -un grupo de peces con mandíbulas del período Devónico, ya extinguidos- presentaban un aspecto similar a las rayas.
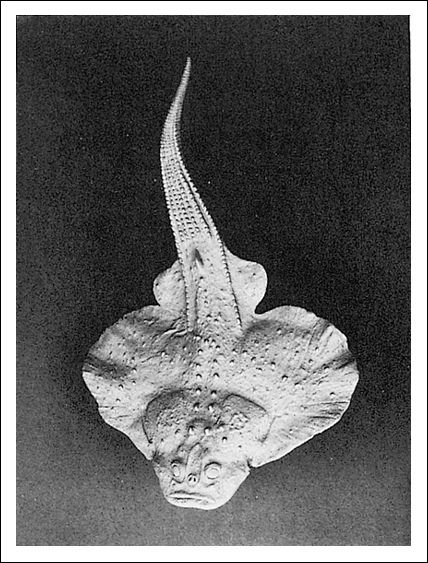
Un placodermo.
Los placodermos fuertemente acorazados constituyeron un grupo de peces mandibulados
muy exitosos en el Devónico. No se ha alcanzado un consenso con respecto
a sus relaciones con otros grupos de peces.
Otro grupo importante de peces
es el de los osteictios, cuyo esqueleto interno en la gran mayoría es
óseo y no cartílaginoso, como ocurre en los condrictios. El grupo
de los osteictios comprende dos subgrupos: los que presentan aletas con rayos
-o actinopterigios-, que incluyen prácticamente a todos los peces de
agua dulce y salada más comunes, y los que presentan aletas carnosas
-o sarcopterigios-, representados en la actualidad por los peces pulmonados,
o dipneustos, de agua dulce.
Mientras que la historia evolutiva
de los condrictios transcurrió principalmente en medio marino, los osteictios
fueron conspicuos habitantes de las aguas continentales al comienzo de su historia.
Muchas formas primitivas de
osteictios parecen haber tenido pulmones -o estructuras análogas a pulmones-
además de branquias. Estos pulmones -que en forma simplificada son sacos
conectados al tubo digestivo- servían como estructuras accesorias de
las branquias y es posible que constituyeran una adaptación especial
que les permitía vivir en aguas poco oxigenadas, principalmente en agua
dulce estancada, pobre en oxígeno,debido a la descomposición de
materia orgánica o al el florecimiento de algas. En algunos de esos linajes
de osteictios, el pulmón pudo haberse transformado en una vejiga aérea
o natatoria. La mayoría de los osteictios vivientes tienen vejigas natatorias
llenas de gas que sirven como cámaras de flotación u órganos
de producción de sonido; algunos de ellos aún conservan una conexión
entre la vejiga y el tracto digestivo. Cuando asciende o desciende en el agua,
el pez puede vencer la gravedad introduciendo gases o expulsándolos de
su vejiga por medio del torrente sanguíneo.
Otro linaje de osteictios está representado actualmente por los peces pulmonados. Estos peces pueden vivir en agua que no tiene suficiente oxígeno y utilizar sus pulmones para captar el oxígeno del aire.

Un pez pulmonado moderno, Protopterus annectens.
Cuando llegan las estaciones secas, los miembros de Protopterus annectensse
introducen en el fango, que termina endureciéndose alrededor de ellos.
Las glándulas de moco debajo de la piel secretan una película
impermeable alrededor del cuerpo, que evita la evaporación. Solamente
la boca queda expuesta. Durante este período, el pez toma una bocanada
de aire sólo aproximadamente una vez cada dos horas.
Un linaje de sarcopterigios
predadores devónicos se adaptó a vivir en aguas poco profundas
explotando los recursos de las áreas marginales de lagos o estuarios.
Estos peces, de cráneos relativamente grandes y gruesas escamas, presentaban
características que eran funcionales en el agua y la tierra, aunque no
de la misma manera. Esto facilitó la transición de los hábitat
acuáticos a los terrestres de los vertebrados. Presumiblemente, estos
peces podían respirar oxígeno atmosférico nadando hacia
la superficie y tragando aire o -en aguas poco profundas- encaramándose
con sus aletas pectorales para elevar sus cabezas hacia la superficie. Los miembros
reforzados pudieron haber sostenido el peso del animal en aguas que no eran
suficientemente profundas para flotar en ellas.
Los anfibios tuvieron su origen
de un grupo de peces óseos sarcopterigios. Los anfibios vivientes incluyen
los anuros -ranas y sapos que carecen de cola después de la metamorfosis-,
las salamandras -que tienen cola durante todo su ciclo vital- y los ápodos
-que carecen de miembros y de vida subterránea-. Su piel fina, habitualmente
sin escamas y que sirve como un órgano respiratorio accesorio, los diferencia
de los reptiles. Los anuros tienen pulmones hacia cuyo interior fuerzan aire
pero algunas salamandras respiran enteramente a través de la piel y de
las membranas mucosas de su garganta. Los anfibios son muy vulnerables a la
desecación, dado que el agua se evapora rápidamente a través
de su piel.
Los anfibios presentan una gran
diversidad de modos reproductivos, mayor que la observada en otros grupos de
vertebrados, especialmente en los amniotas. En cada uno de los tres grupos vivientes
de anfibios hay tendencias hacia la terrestrialidad. La mayoría de los
anuros tienen dos fases en su ciclo vital: una en el agua y otra en la tierra
(de allí su nombre, de amphi y bios que significa "doble vida").
Prácticamente todos los anuros tienen fertilización externa. Los
óvulos se depositan en el agua, o cerca del agua, y allí se fecundan.
Nacen como larvas acuáticas con branquias (renacuajos) que, luego de
profundos cambios dan lugar a adultos con respiración pulmonar. Los adultos
pueden vivir fuera del agua, aunque hay muchas variaciones sobre este tema.
Los ápodos y la mayoría de las salamandras tienen fecundación
interna. Entre estas últimas, los machos depositan paquetes de espermatozoides,
ya sea en el agua o en el suelo húmedo, que son recogidos por las hembras.
Muchos anfibios vivientes omiten completamente la etapa larval de vida libre.
Los huevos, que pueden depositarse en tierra, en un tronco hueco o en una hoja
acopada, o inclusive pueden ser transportados por el padre, nacen como una versión
en miniatura del adulto.
La aparición del huevo
amniota en un grupo de tetrápodos posibilitó la conquista más
completa de los hábitat terrestres. Este grupo de tetrápodos -denominados
amniotas- está representado actualmente por los reptiles, aves y mamíferos;
su origen se remonta al Carbonífero.
El huevo amniota -o cleidoico- se caracteriza por tener una cáscara flexible o rígida que provee protección mecánica, pero que al mismo tiempo permite el paso de gases respiratorios y vapor de agua a través de ella. La albúmina (clara del huevo) constituye un reservorio de agua y proteínas, así como la yema. Pero, lo que realmente distingue este tipo de huevos, es la presencia de tres membranas extraembrionarias: amnios y corion (que rodean al embrión) y alantoides, que funciona como reservorio de los desechos nitrogenados producidos por el metabolismo del embrión y que, en consecuencia, no necesitan ser reprocesados.
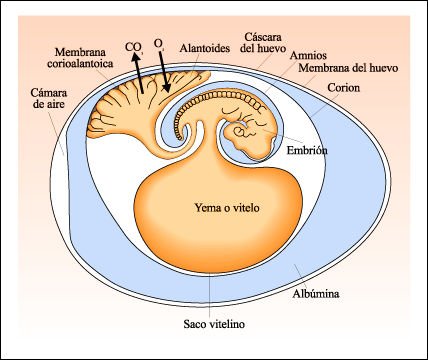
El huevo amniota.
Las membranas del huevo amniota, producidas como excrecencias del embrión
a medida que se desarrolla, rodean y protegen al embrión y a la yema
(su fuente de alimento). La cáscara y la membrana del huevo, que son
impermeables al agua, pero permeables a los gases, se agregan al embrión
cuando éste desciende por el tracto reproductivo materno.
Los reptiles tienen una piel
fuertemente queratinizada, habitualmente cubierta por escamas protectoras, y
con pocas glándulas. Esto posibilita su existencia terrestre y su actividad
diurna. Los reptiles vivientes incluyen lagartos, serpientes, tortugas y cocodrilos.
Durante toda la prolongada era
Mesozoica -unos 150 millones de años- los dinosaurios dominaron la vida
terrestre. Luego, hace aproximadamente 65 millones de años, desaparecieron,
dejando sólo una única línea de descendientes, las aves.
La causa de su extinción ha sido tema de especulación desde que
los primeros fósiles de dinosaurios se descubrieron en el siglo XIX,
pero sólo recientemente han aparecido nuevos datos que permitieron formular
hipótesis comprobables.
Las aves son, esencialmente,
reptiles especializados en el vuelo. Sus cuerpos contienen sacos aéreos
y sus huesos son huecos.
El hueso más masivo del
esqueleto de las aves es la quilla o esternón, en la que se insertan
los músculos que mueven las alas. Las aves voladoras presentan una gran
reducción en el peso; el sistema reproductor de las hembras se ha reducido
a un solo ovario, y aun éste se hace bastante grande como para ser funcional
sólo en la época de apareamiento.
Las aves tienen plumas, que
son su característica morfológica más notable. Son endotérmicas;
mantienen una temperatura corporal alta y constante gracias a la alta tasa metabólica
y a la excelente aislación provista por las plumas. Esos dos factores
son ventajosos en conjunción pero las plumas, como veremos, podrían
haber resultado inicialmente de la selección para una función
distinta de la termorregulación. Sólo los animales endotérmicos
requieren de aislación; la aislación sería una desventaja
para los animales que calientan sus cuerpos exponiéndolos al ambiente.
Las aves también tienen escamas en sus patas, un relicto de su ascendencia.
Muchas aves nacen en un estadio muy inmaduro y virtualmente todas requieren
un largo período de cuidados maternos.
Los primeros mamíferos,
que aparecieron en la etapa más temprana del Mesozoico, eran pequeños
y probablemente nocturnos; esto les permitió evitar a los dinosaurios
carnívoros que estaban activos durante las horas de luz. Durante casi
todo el Mesozoico, los dinosaurios fueron los vertebrados terrestres más
conspicuos, y su desaparición dejó un vacío que permitió
la radiación mamaliana. Sin embargo, a pesar de la rápida diversificación
de los mamíferos y el notable incremento de tamaño en muchos linajes
a comienzos del Terciario, éstos no alcanzaron inmediatamente la amplitud
ecológica ni la talla de los dinosaurios.
Los mamíferos presentan
una serie de caracteres anatómico-fisiológicos que los distinguen
de otros vertebrados. Entre ellos, pueden mencionarse los siguientes: l) tienen
pelo, 2) mantienen una alta temperatura corporal generando calor metabólicamente,
3) presentan articulación mandibular entre el hueso dentario de la mandíbula
inferior y el hueso escamoso de la superior, 4) presentan heterodoncia -dientes
altamente diferenciados en distintas partes de la boca- con precisión
oclusal (los dientes de arriba y de abajo tienen encastre preciso) y sólo
dos generaciones dentarias, 5) suministran leche a su progenie por glándulas
especializadas (glándulas mamarias).
Casi todas las especies de mamíferos son vivíparas. Sin embargo, algunos mamíferos como los monotremas, que incluye al ornitorrinco, depositan huevos con cáscara, pero nutren a su progenie después del nacimiento. En cambio, los marsupiales, que incluyen a las comadrejas y a los canguros, entre otros, son vivíparos. Sin embargo, difieren de los placentarios en que, aunque frecuentemente se forma una placenta con el saco vitelino, la gestación es muy breve y las crías nacen en una condición extremadamente inmadura; a menudo, las mantienen en una bolsa especial donde se amamantan y continúan su desarrollo. La mayoría de los mamíferos comunes son placentarios, llamados así porque presentan una conexión materno-embrionaria denominada placenta. La placenta es una estructura vascular derivada de las membranas extraembrionarias del huevo amniota y el útero materno, que permite la alimentación del embrión a través de la sangre materna. Como resultado, la progenie se desarrolla hasta una etapa mucho más avanzada antes del nacimiento. Así, las crías tienen protección y alimento durante su período más vulnerable.
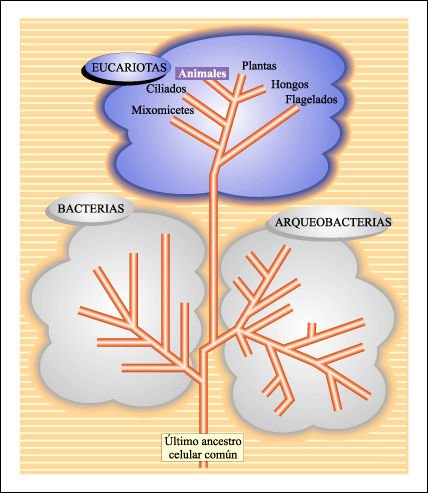
Esquema global de las relaciones evolutivas de los seres vivos.
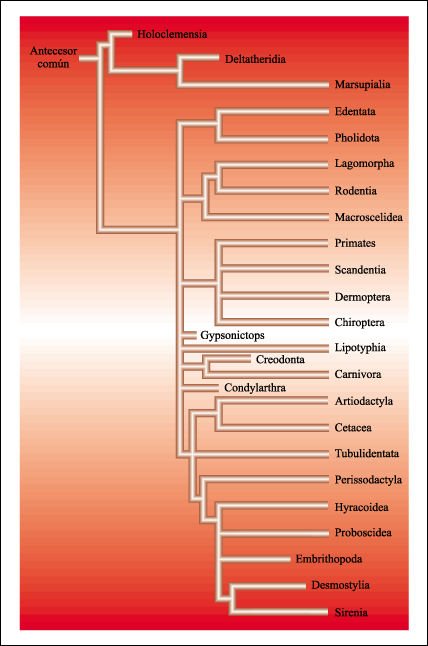
En esta figura se muestran las relaciones tentativas de diversos grupos de mamíferos.
Los primates, el grupo al cual pertenecemos, son mamíferos placentarios
que retienen las cuatro clases de dientes (caninos, incisivos, premolares y
molares), los primeros dígitos oponibles (pulgares y, a veces, el dedo
grande de los pies), garras transformadas en uñas planas, dos glándulas
mamarias en el pecho (y no en el abdomen), ojos dirigidos frontalmente, y un
cerebro relativamente grande con una corteza cerebral con circunvoluciones.
Muchas de estos caracteres básicos de los primates permiten la subsistencia
en hábitat arbóreos especializados: localización visual
y captura manual de insectos arborícolas como presas.
Nos distinguimos de otros primates
por nuestra postura erecta, piernas largas y brazos cortos, frente alta y una
mandíbula pequeña y escaso pelo corporal.


