
Capítulo 35.
Las plantas
con flores: Introducción
En la actualidad, las plantas más abundantes de la Tierra, que evolucionaron
más recientemente, son las angiospermas.
Las angiospermas se caracterizan
por estructuras reproductoras especializadas, las flores, en las cuales ocurre
la reproducción sexual. Para estas plantas, un nuevo ciclo de vida comienza
cuando un grano de polen, portado por el viento o frecuentemente desprendido
del cuerpo de un insecto forrajero, entra en contacto con el estigma de una
flor de la misma especie.
Después de la fecundación,
el cigoto se divide mitóticamente y forma el embrión. A medida
que el embrión crece, sus células comienzan un proceso de diferenciación.
La semilla de las angiospermas consiste en el embrión, la cubierta de
la semilla y el alimento almacenado; el fruto se desarrolla de la pared del
ovario. Cuando el ovario madura en fruto y se forman las semillas, los pétalos,
estambres y otras partes de la flor generalmente caen.
No siempre las semillas de las
plantas encuentran las condiciones más propicias para su desarrollo.
En el transcurso de su evolución, se han seleccionado las características
que permitieron a las plantas resistir condiciones adversas. La principal de
estas adaptaciones es, posiblemente, la capacidad para permanecer latentes.
Reproducción sexual:
la flor
La mayoría de las flores consisten en cuatro conjuntos de piezas: sépalos, pétalos, estambres y carpelos. Se piensa que cada pieza floral es, evolutivamente hablando, una hoja modificada. En las monocotiledóneas, cada una de las piezas florales habitualmente está presente en múltiplos de 3; en las dicotiledóneas, por el contrario, las piezas florales típicamente aparecen en múltiplos de 4 ó 5.
Flor de un peral.
Ésta es una flor completa,
lo que significa que contiene las cuatro partes florales: sépalos, pétalos,
estambres y carpelos. Es también una flor perfecta, lo que significa
que contiene tanto a las estructuras masculinas (estambres) como femeninas (carpelos).
Cada estambre consiste en una antera con polen y su filamento. Cada carpelo
está formado por un estigma, un estilo y un ovario.
Hay diversos tipos de flores. Una flor que contiene estambres y carpelos se conoce como flor perfecta. Sin embargo, en algunas especies, las flores son imperfectas, o sea, son masculinas (estaminadas) o femeninas (carpeladas). En la misma planta, pueden estar presentes flores masculinas y femeninas; se dice que estas plantas son monoicas. Las especies en las cuales las flores masculinas y femeninas se encuentran en plantas separadas se conocen como dioicas.

Flores: formas y variaciones.
a) Una magnolia, Magnolia virgilniana.
Los carpelos forman un receptáculo en forma de cono, del cual surgen
los estilos curvos. Los estambres de color crema, algunos de los cuales han
caído, rodean a los carpelos. Las magnolias son flores muy primitivas;
tienen numerosas piezas florales dispuestas en un patrón espiralado.
b) La orquídea sandalia de dama, Paphiopedilum invincible . Las orquídeas
son flores altamente especializadas, cuyos estilos y filamentos están
fusionados en una sola estructura que lleva el estigma y, a corta distancia,
una antera. El "labio" de la flor es un pétalo modificado que
puede servir como "plataforma de aterrizaje" para los insectos. Las
Orchidaceae es la segunda familia más grande de las plantas con flor.
c) El girasol, Helianthus annuus ("flor anual del Sol"). En el girasol
y otras compuestas, numerosas flores individuales constituyen lo que se denomina
capítulo, que actúa como una sola flor enorme que atrae a los
insectos. La porción central del capítulo está formada
por varias flores pequeñas (flores del dico o flósculos) separadas.
Cada uno de estos flósculos consta de un par de carpelos fusionados -que
forman un ovario único- y anteras fusionadas encerradas en una pequeña
corola de pétalos también fusionados. Esta porción central
está rodeada por flores radiadas (de pétalos amarillos) que frecuentemente
son estériles. Las Compuestas (familia Asteraceae), es la familia más
numerosa entre las plantas con flor. d) En Hibiscus, una columna de estambres
está fusionada alrededor del estilo. e) Estambre y estigma de un crocus
púrpura. f) Maíz (Zea mays), una especie monoica. Las flores masculinas
y femeninas separadas son llevadas en la misma planta. Las espiguillas, en la
parte superior del tallo, son conjuntos de flores masculinas (productoras de
polen). Cada hebra de "seda" o "barba" que se ve surgir
de la espiga de maiz, en la parte inferior, es el estigma y el estilo combinados
de la flor femenina. g) Cicuta acuática, miembro de la familia de la
zanahoria, constituye otro ejemplo de inflorescencia en el cual varias flores
se encuentran ubicadas laxamente sobre una misma rama floral.
Cuando el grano de polen se
libera de la antera, habitualmente contiene tres núcleos haploides: dos
núcleos espermáticos o generativos y un núcleo del tubo
o vegetativo. Todos están rodeados por la pared externa gruesa del grano
de polen. El grano de polen contiene sus propios nutrientes y tiene una cubierta
externa muy dura que lo protege.
Una vez en el estigma, el grano
de polen germina y, bajo la influencia del núcleo del tubo, crece el
tubo polínico a través del estilo hasta penetrar en un óvulo.
Cada óvulo contiene un gametofito femenino. En muchas especies, el gametofito femenino está formado por siete células, con un total de ocho núcleos haploides. Una de las siete células es la ovocélula u oósfera, que contiene un único núcleo haploide. A cada lado de la ovocélula hay una pequeña célula conocida como sinérgida. En el extremo opuesto del gametofito hay tres células pequeñas, las células antípodas, cuya función, si es que la tienen, se desconoce. La célula central grande contiene dos núcleos haploides, llamados núcleos polares.
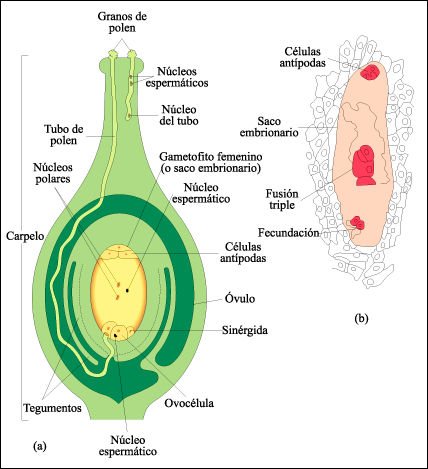
Fecundación en las angiospermas.
El tubo de polen del gametofito
masculino, o grano de polen, crece a través del estilo y entra en un
óvulo que contiene el gametofito femenino de siete células (el
saco embrionario). Uno de los núcleos espermáticos se une con
la ovocélula, formando el cigoto. El otro núcleo espermático
se fusiona con los dos núcleos polares contenidos en una sola célula
grande (que en el dibujo ocupa la mayor parte del óvulo). Esta fusión
triple produce una célula triploide (3n) de la cual se originará
el endosperma. El carpelo que se muestra aquí contiene un solo óvulo.
El endosperma rodea y nutre al embrión en desarrollo. Estos fenómenos
extraordinarios de fecundación y fusión triple, llamados en conjunto
doble fecundación ocurren, entre todos los seres vivientes, sólo
en las plantas con flor.
El embrión
A medida que el embrión
crece, sus células comienzan un proceso de diferenciación. Las
dos células que surgen de la primera división del cigoto establecen
una polaridad en el embrión. La célula inferior normalmente da
una estructura multicelular llamada suspensor y la célula superior se
convierte en el embrión verdadero.
En las primeras etapas, el embrión
consiste en una masa globular de células sobre el suspensor. Las células
del suspensor intervienen activamente en el envío de nutrientes al embrión.
A medida que procede el desarrollo del embrión, los cambios en su estructura interna dan como resultado la formación de tres tejidos embrionarios distintos conocidos como meristemas primarios. Simultáneamente, o un poco después, ocurre la aparición de un cotiledón ("hoja seminal") en las monocotiledóneas o de dos cotiledones en las dicotiledóneas. Gradualmente, el embrión adquiere su forma característica.
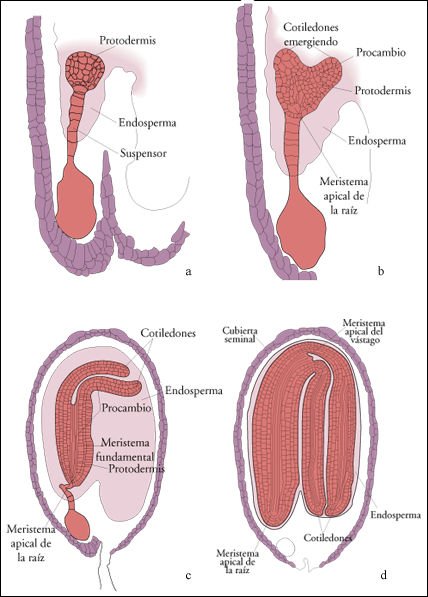
Algunas etapas en el desarrollo del embrión del morral del pastor (Capsella bursa-pastoris), una dicotiledónea.
a) El primer tejido embrionario
que se diferencia es la protodermis, a partir de la cual se desarrollará
la cubierta externa de la planta recién nacida. La célula grande
en la parte inferior del embrión es la célula basal del suspensor.
b) Los cotiledones ("hojas seminales") comienzan a emerger. El procambio,
un segundo tejido embrionario, luego originará los tejidos vasculares
de la planta. c) Los cotiledones se han desarrollado más. Una diferenciación
adicional ha producido un tercer tejido embrionario, el meristema fundamental,
del cual derivará la masa de tejido de las hojas, tallos y raíces
jóvenes. Los tres tejidos embrionarios, conocidos como meristemas primarios,
tienen continuidad entre los cotiledones y el eje del embrión. d) El
embrión maduro dentro de su cubierta seminal protectora. Los meristemas
apicales de la raíz y del vástago están claramente diferenciados.
En las primeras etapas del crecimiento
embrionario ocurren divisiones celulares en todo el cuerpo de la planta joven.
A medida que el embrión se desarrolla, la adición de nuevas células
se restringe gradualmente a ciertas partes del cuerpo de la planta: los meristemas
apicales, localizados cerca del ápice de la raíz y del vástago.
Durante el resto de la vida de la planta, el crecimiento primario -que implica
principalmente el alargamiento del cuerpo de la planta- se origina en los meristemas
apicales de las raíces y vástagos.
La semilla y el fruto
En las angiospermas, la semilla consiste en el embrión -que se desarrolla a partir de la ovocélula fecundada-, la reserva de alimento -que consiste en el endosperma o deriva de éste- y la cubierta de la semilla -que se desarrolla de la capa o capas más externas del óvulo -. Al mismo tiempo, el fruto se desarrolla de la pared del ovario (la base del carpelo). Cuando el ovario madura en fruto y se forman las semillas, los pétalos, estambres y otras partes de la flor generalmente caen.
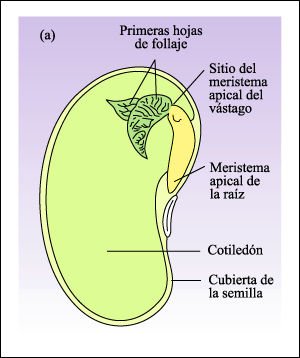
Semillas. a) Dicotiledónea.
En las dicotiledóneas como el guisante común, el endosperma es digerido a medida que el embrión crece y la reserva de alimento se transfiere a los cotiledones carnosos.
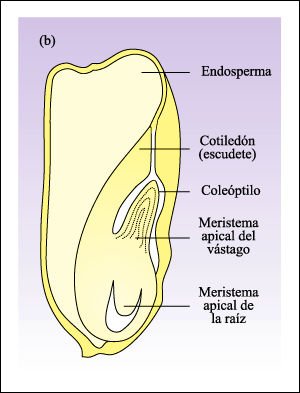
b) Semilla de maíz.
En el maíz y otras monocotiledóneas,
el cotiledón único, conocido en el maíz como escudete,
absorbe las reservas alimenticias del endosperma. El coleóptilo es una
vaina que encierra al meristema apical del vástago; es la primera estructura
que aparece por encima del suelo después de que la semilla germina.
Los frutos adoptan formas diferentes. Muchas de estas formas son adaptaciones a una variedad de mecanismos de dispersión. Los frutos generalmente se clasifican en simples, agregados o múltiples, según la disposición de los carpelos en la flor.
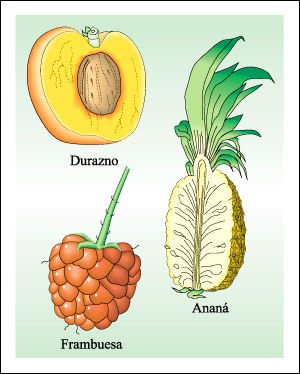
Un fruto simple, un fruto agregado y, a la derecha, un fruto múltiple.
Los frutos simples son los más diversos. Cuando maduran pueden ser blandos
y carnosos o secos. Los tres tipos principales de frutos carnosos son la baya,
la drupa y el pomo.
Los frutos simples secos se clasifican en dos grandes grupos, dehiscentes e indehiscentes. Los frutos dehiscentes maduros se abren y liberan las semillas cuando están todavía unidos a la planta materna; por oposición, las semillas de los frutos indehiscentes se encuentran dentro del fruto cuando éste se desprende de la planta.
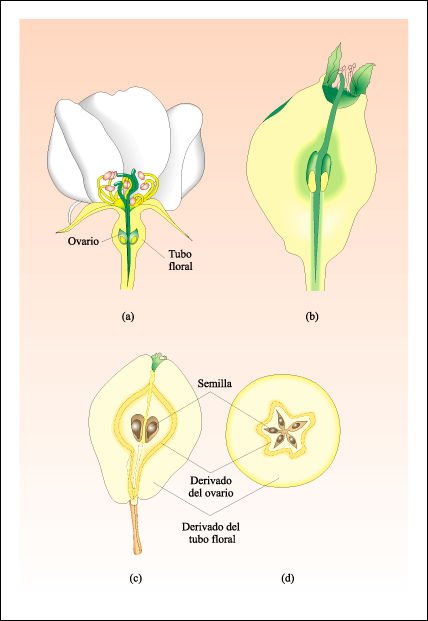
Desarrollo y estructura de la pera, un pomo.
a) Flor del peral. El ovario es la porción basal del carpelo. b) Flor
más vieja, después de que han caído Los pétalos.
c) Corte longitudinal y d) corte transversal del fruto maduro. El corazón
de la pera es la pared del ovario maduro. La parte carnosa comestible se desarrolla
a partir del tubo floral.
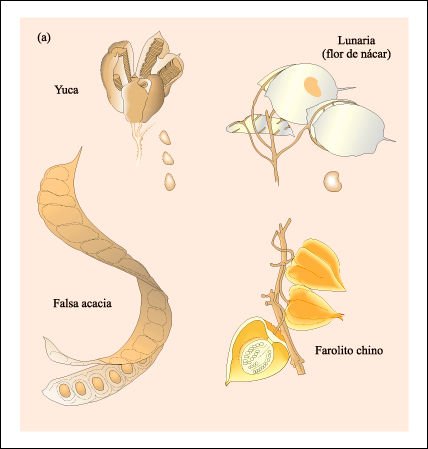
Ejemplos de frutos. a) Frutos dehiscentes
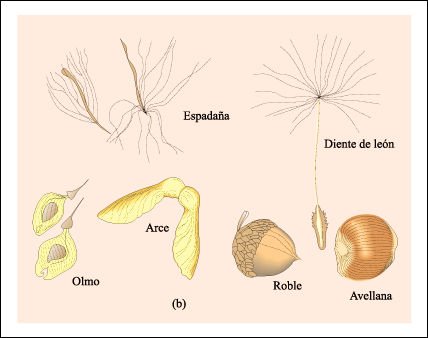
b) Frutos indehiscentes.
La distancia a la que son dispersadas
las semillas de los frutos dehiscentes desde la planta madre frecuentemente
depende de la fuerza con que el fruto se abre. Muchos frutos indehiscentes son
llevados a una gran distancia de la planta madre, ya sea por el viento o por
animales que los ingieren, antes de que se liberen las semillas.
Adaptaciones a los cambios estacionales
La latencia, estado en el que
las plantas pueden soportar condiciones ambientales rigurosas, permite a las
angiospermas resistir los períodos de sequía o de frío
inadecuados para el crecimiento vegetal. Las angiospermas se clasifican en anuales,
bienales y perennes, dependiendo de si el cuerpo de la planta muere al final
de una estación de crecimiento (anual), después de dos estaciones
(bienal), o si las porciones vegetativas del cuerpo de la planta persisten año
tras año (perenne).
Durante el desarrollo, las semillas
adquieren resistencia a la desecación y cuando están maduras se
encuentran en estado de latencia. Algunas especies tienen semillas que no toleran
la deshidratación; se las denomina recalcitrantes. Las semillas latentes
se encuentran listas para la germinación, pero necesitan una combinación
adecuada de factores externos para poner en marcha la maquinaria metabólica
de la semilla.
Las semillas maduras de muchas
plantas no germinan inmediatamente después de su dispersión, aunque
encuentren condiciones externas favorables, debido a restricciones internas
para la germinación. En esos casos, se dice que las semillas se encuentran
en estado de dormición. La dormición puede estar impuesta tanto
por los tejidos que rodean al embrión, por causas inherentes al mismo
tejido o por una combinación de ambos factores.
Una vez que se produce la ruptura
de la dormición, las semillas entran en estado de latencia en el cual,
si las condiciones externas son las apropiadas, se producirá la germinación.
La ruptura de la dormición y su relación con factores ambientales
permite que la germinación ocurra cuando las condiciones para el establecimiento
exitoso de las plántulas sea más favorable.


