Capítulo 36.
El cuerpo
de la planta y su desarrollo
Las plantas son organismos fotosintéticos multicelulares adaptados a
vivir en tierra firme. El cuerpo de la planta tiene áreas fotosintéticas
especializadas llamadas hojas, estructuras de conducción y de soporte,
los tallos, y órganos que fijan la planta al suelo y absorben agua y
nutrientes esenciales de él, las raíces.
Cuando una semilla germina,
el crecimiento de la raíz y del vástago procede de los meristemas
apicales del embrión. Ciertas células dentro del meristema no
pierden la capacidad de dividirse. Otras se alargan y luego se diferencian formando,
según su posición, las distintas células especializadas
de la planta. Estas células, en combinaciones varias, forman los tres
sistemas de tejidos -dérmico, fundamental y vascular- que son continuos
en todo el cuerpo de la planta. Las células parenquimáticas, de
paredes delgadas y multilaterales, son las células más comunes
de las plantas.
Las áreas fotósinteticas
por excelencia de una planta son las hojas. La estructura de una hoja parece
resultar de un compromiso entre tres presiones evolutivas opuestas: exponer
una superficie fotosintética máxima a la luz, conservar agua y,
al mismo tiempo, asegurar el intercambio de gases necesarios para la fotosíntesis.
Las plantas, a diferencia de
los animales, continúan creciendo durante todo su ciclo de vida. La raíz
embrionaria es la primera estructura que aparece de la semilla al iniciarse
la germinación. Las raíces fijan la planta al suelo e incorporan
agua y minerales esenciales. El crecimiento de las raíces necesita de
una gran inversión de energía por parte de las plantas; esta energía
proviene de los enlaces químicos de las moléculas carbonadas que
circulan por el vástago.
Los tallos son las estructuras
de conducción y de soporte de las plantas. También pueden estar
adaptados al almacenamiento de alimento o de agua.
La altura (o longitud) de las
partes aéreas de la planta se incrementa por el crecimiento primario
del sistema de vástago. El sistema del vástago está formado
por el tallo y otras estructuras que se desarrollan a partir de él.
El crecimiento secundario es
el proceso por el cual las plantas leñosas incrementan su diámetro.
Las células y los tejidos
del cuerpo vegetal
Las células del embrión
de una angiosperma se diferencian tempranamente en su desarrollo en tres tejidos
distintos: la protodermis, el procambio y el meristema fundamental. Estos tejidos
embrionarios, conocidos como meristemas primarios, producen los tres sistemas
de tejidos que son continuos en todo el cuerpo de la planta. La protodermis,
el primer tejido que se diferencia, es el origen del sis-tema de tejido dérmico,
que proporciona una cubierta externa protectora para todo el cuerpo de la planta.
El procambio, el tejido que se diferencia luego, origina el sistema de tejido
vascular, compuesto por xilema y floema. El xilema transporta agua y minerales
disueltos, mientras que el floema transporta azúcares disueltos y otros
compuestos or-gánicos desde las células fotosintéticas
(autótrofas) de las hojas y de los tallos verdes a las células
no fotosintéticas (heterótrofas) de la planta. Los tejidos vasculares
están rodeados por el sistema de tejido fundamental, derivado del meristema
embrionario fundamental. Las principales diferencias en la estructura de las
hojas, tallos y raíces reside en la distribución relativa de los
sistemas de tejido vascular y fundamental.
Las células que se encuentran
más frecuentemente en el cuerpo de la planta son del tipo conocido como
parénquima. Estas células, que aparecen en los tres sistemas de
tejidos y predominan en los tejidos fundamentales, son típicamente poliédricas
y tienen paredes delgadas y flexibles. Además de un núcleo, mitocondrias
y otras organelas, las células parenquimáticas generalmente contienen
plástidos que, según la ubicación de la célula,
pueden ser cloroplastos, leucoplastos o cromoplastos.
Además de la fotosíntesis,
las células parenquimáticas desempeñan una variedad de
funciones esenciales en la planta, que incluyen la respiración y el almacenamiento
de alimento y agua. Cada uno de los sistemas de tejidos contienen también
tipos celulares adicionales, especializados en funciones particulares de cada
órgano.
Hojas
Las hojas son las áreas
fotosintéticas de la planta. Las células fotosintéticas
de las hojas son células parenquimáticas que forman dos tipos
de tejidos: parénquima en empalizada, constituido por células
alargadas y densamente empaquetadas ubicadas justo por debajo de la superficie
superior de la hoja, y parénquima esponjoso, que consiste en células
de contorno irregular situadas en el interior de la hoja y con grandes espacios
intercelulares. Estos espacios están llenos de gases, que incluyen vapor
de agua, oxígeno y dióxido de carbono. La mayor parte de la fotosíntesis
ocurre en las células en empalizada, que están especializadas
en la captación de la luz.
El parénquima en empalizada
y el parénquima esponjoso constituyen el tejido fundamental de la hoja,
conocido como mesófilo. El mesófilo está envuelto casi
herméticamente por las células epidérmicas, que secretan
una sustancia cérea llamada cutina que forma una cubierta, la cutícula,
sobre la superficie externa de la epidermis. Las células epidérmicas
y la cutícula son transparentes, lo que permite que la luz las atraviese
y penetre en las células fotosintéticas.
Las sustancias entran y salen de las hojas a través de dos estructuras completamente diferentes: los haces vasculares y los estomas. El agua y los minerales disueltos son transportados a las hojas, y los productos de la fotosíntesis son transportados fuera de ellas, por medio de los haces vasculares. Los haces vasculares atraviesan los pecíolos y se continúan con los tejidos vasculares del tallo y la raíz.
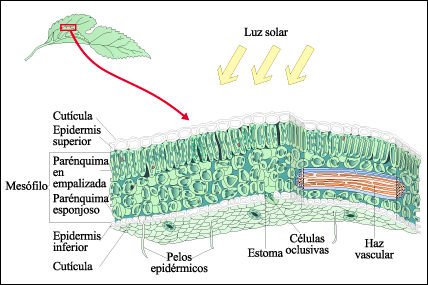
Estructura de una hoja.
La fotosíntesis ocurre en las células denominadas en empalizada
y, en menor grado, en el tejido llamado parénquima esponjoso. Los cloroplastos
se indican en verde oscuro. Nótese que el citoplasma, que contiene a
los cloroplastos, está concentrado cerca de la superficie celular y en
la parte central de las células se encuentran vacuolas grandes. Los cloroplastos
se mueven dentro del citoplasma, orientándose hacia el Sol. Los haces
vasculares llevan agua y solutos hacia y desde las células del mesófilo.
El interior de la hoja está encerrado por células epidérmicas
cubiertas por una capa cérea, la cutícula. Las aberturas en la
epidermis son los estomas que permiten el intercambio de gases. Las células
oclusivas que rodean a los estomas también tienen cloroplastos
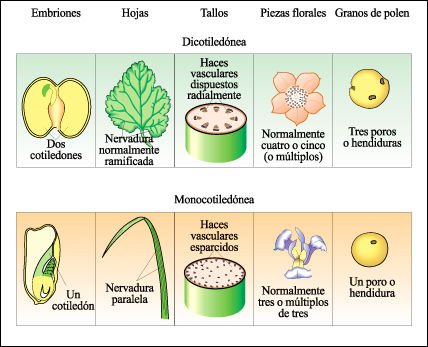
Las dos clases de angiospermas son las dicotiledóneas y las monocotiledóneas.
Los nombres dicotiledóneas y monocotiledóneas se refieren al hecho
de que el embrión de una dicotiledónea tiene dos cotiledones (hojas
seminales) y el embrión de una monocotiledónea tiene uno. Otras
diferencias características se hacen visibles en el cuerpo de la planta.
Los haces vasculares principales de las hojas de las dicotiledóneas habitualmente
se disponen en red; los haces vasculares de las hojas de las monocotiledóneas
habitualmente son paralelos. En los tallos de las dicotiledóneas, los
haces de te-jido vascular se disponen alrededor de un núcleo central
de tejido fundamental; en los tallos de las monocotiledóneas, los haces
vasculares están dispersos en el tejido fundamental. Existen también
diferencias características en el número de piezas florales y
en el número de hendiduras de los granos de polen entre ambas clases
de angiospermas.Los gases -oxígeno y dióxido de carbono- entran
y salen de las hojas por difusión a través de estomas.
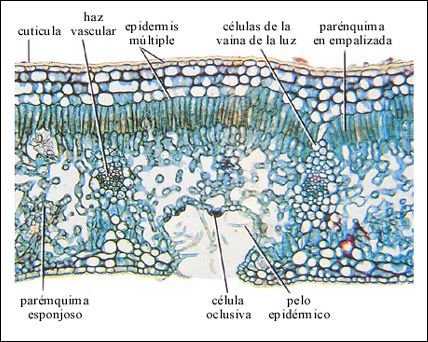
Corte transversal de la hoja de una adelfa o laurel rosa, una angiosperma adaptada a climas secos.
En la parte superior, una cutícula muy gruesa cubre la epidermis múltiple,
así llamada porque consta de cuatro capas de células. El estoma
se encuentra dentro de una cripta estomática, tapizada con pelos epidérmicos.
Las hojas presentan una variedad de formas y tamaños, que van desde frondes grandes a escamas diminutas. Estas diferencias en la morfología y tamaño guardan una estrecha relación con los ambientes en los cuales vive la planta.

Hojas modificadas.
a) Espinas de un nopal, o chumbera, cacto gigante fotografiado en las Islas
Galápagos. b) Hojas suculentas adaptadas al almacenamiento de agua (Sedum).
c) Zarcillo de una planta de guisante. En la planta de guisante, que tiene hojas
compuestas, sólo los folíolos individuales se modifican como zarcillos;
otros folíolos de hojas compuestas son aplanados, lo que proporciona
una superficie amplia para la fotosíntesis.
Características
del crecimiento de la planta
Los embriones de muchas angiospermas pasan por un período de latencia antes de que germine la semilla. Con la germinación se reinicia el crecimiento, se rompe la cubierta de la semilla y surge el esporofito joven. Las primeras hojas de follaje se abren hacia el Sol y comienzan a hacer fotosíntesis, mientras que, internamente, continúa el proceso de crecimiento que da origen al cuerpo de la planta. En los primeros estadios del desarrollo, el crecimiento del joven esporofito depende de las reservas acumuladas en la semilla.
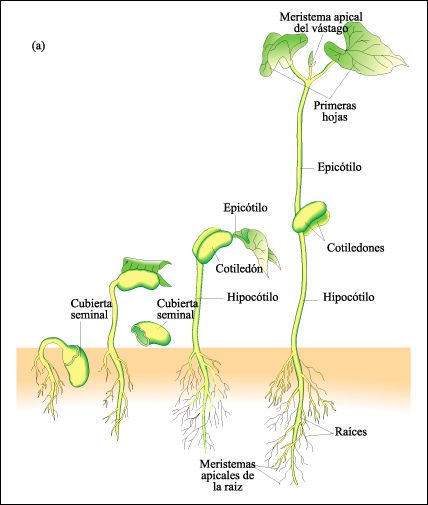
a) Desarrollo de una plántula de frijol, una dicotiledónea . x
Antes de la germinación, la semilla absorbe agua y se hincha, rompiendo
la cubierta seminal. Primero emerge la raíz joven, seguida del hipocótilo
("debajo de los cotiledones"). Los cotiledones finalmente se marchitarán
y caerán. Luego emerge el epicótilo que se encuentra entre cotiledón
y haz de hojas.

b) Desarrollo de una plántula de maíz, una monocotiledónea .
La primera estruc-tura que aparece por encima del suelo es el coleóptilo,
que forma una vaina cilíndrica sobre el vástago en crecimiento
de la planta. Típicamente, el resto de endosperma, con el escudete (el
cotiledón único) en su interior, está presente en la joven
plántula.
El
crecimiento primario de la planta implica la diferenciación de los tres
sistemas de tejido, el alargamiento de las raíces y tallos, y la formación
de las raíces laterales y de las ramas. Después de completarse
el desarrollo del embrión, el crecimiento primario posterior se origina
en los meristemas apicales de la raíz y del vástago. Las plantas,
a diferencia de los animales, continúan creciendo durante todo su ciclo
de vida.
Raíces
Las
raíces son estructuras especializadas que fijan la planta al suelo e
incorporan agua y minerales esenciales. La raíz embrionaria, o radícula,
es la primera estructura que rompe la cubierta seminal y se elonga rápidamente.
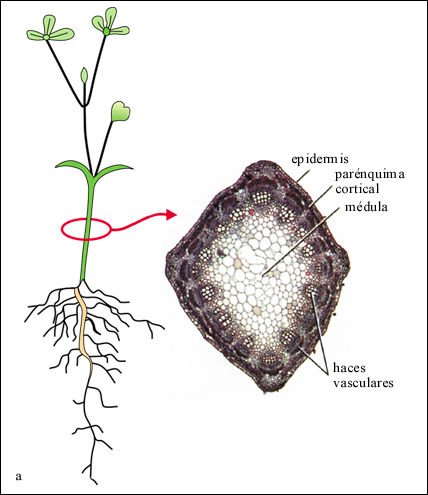
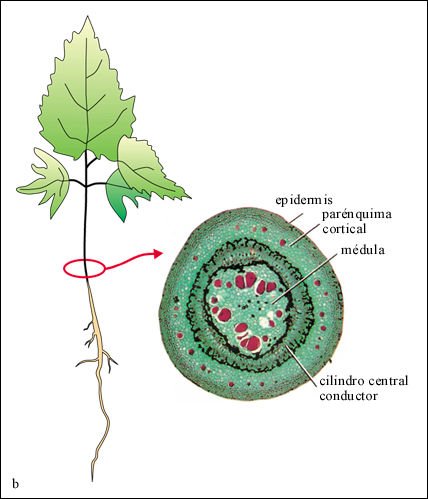
La estructura interna de la raíz de las angiospermas es comparativamente simple. En las dicotiledóneas y en la mayoría de las monocotiledóneas los tres sistemas de tejidos (dérmico, fundamental y vascular) están dispuestos en tres capas concéntricas: la epidermis, la corteza y el cilindro central.
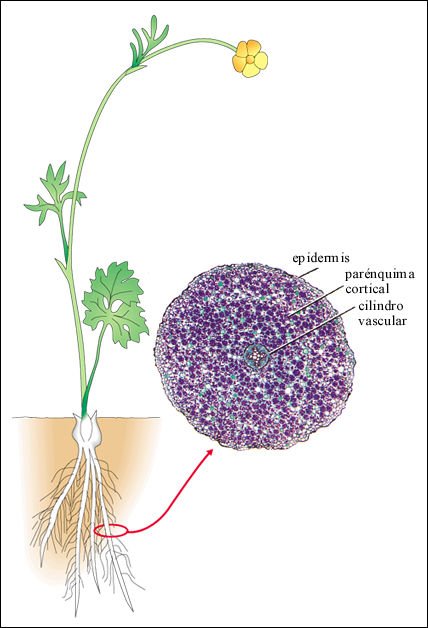
Raíz de un ranúnculo, una dicotiledónea, en corte transversal.
Los
plástidos en las células parenquimáticas de la corteza
contienen granos de almidón, teñidos en púrpura en esta
preparación.
Las raíces jóvenes tienen una capa externa de epidermis y, a lo sumo, una cutícula muy delgada. Prolongaciones de las células epidérmicas forman los pelos radicales, que incrementan en sumo grado la superficie absorbente de la raíz. Debajo de la epidermis está el tejido fundamental de la raíz, la corteza, compuesta principalmente por células parenquimáticas, frecuentemente especializadas en el almacenamiento. La capa más interna de la corteza es la endodermis, una sola capa de células especializadas cuyas pare-des contiguas, en las que se ha depositado suberina, conforman la banda de Caspary.
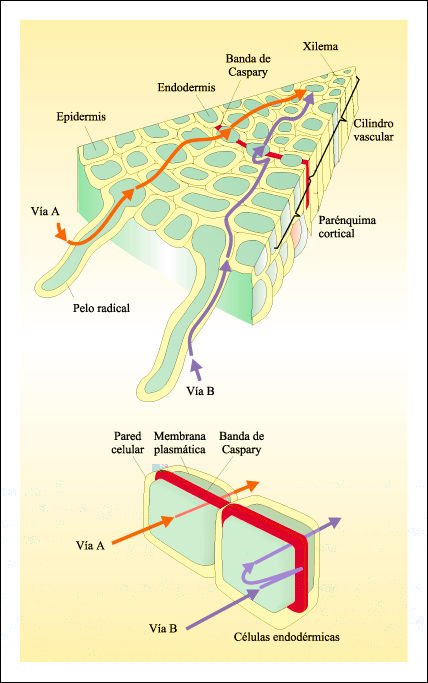
Diagrama de un corte transversal de una raíz que muestra las dos vías de absorción de agua y sustancias disueltas.
La mayor parte de los solutos y parte del agua que entra en la raíz siguen
la vía A, que se indica en color rojo; los solutos penetran a las células
de la epidermis de la raíz por transporte activo o por difusión
facilitada; en tanto que el agua se mueve por diferencia de gradiente de potencial
hídrico. Luego de atravesar las membranas celulares, ya en el simplasma
de la raíz, el agua y los solutos se desplazan de una a otra célula
a través de los plasmodesmos. El simplasma es la continuidad del protoplasma
a través de los plasmodesmos. Otra parte del agua y algunos de los solutos
que entran a la raíz siguen la vía B, indicada en color azul,
moviéndose a través de las paredes celulares y a lo largo de sus
superficies. Nótese, sin embargo, la ubicación de la banda de
Caspary y de qué manera bloquea la vía B en torno a todo el cilindro
vascular de la raíz. Para atravesar la banda de Caspary, tanto el agua
como los solutos tienen que ser transportados a través de las membranas
celulares de las células endodérmicas o de otras células
situadas más externamente, a través de la vía A. Después
que el agua y los solutos han cruzado la endodermis, la mayoria de los solutos
continúan a lo largo de la vía A hasta las células conductoras
del xilema, y la mayor parte del agua retorna a la vía B para cubrir
la distancia que resta hasta las células del xilema. En las zonas más
jovenes de la raíz, donde aún no se ha formado la banda de Caspary,
el agua y los nutrientes esenciales pueden alcanzar el xilema a través
de la vía B.
Inmediatamente por dentro de la endodermis hay otra capa de células, el periciclo, del cual surgen las ramificaciones de la raíz. Por dentro del periciclo están el xilema y el floema.
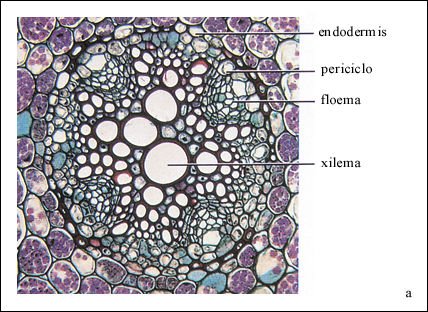
Detalle del cilindro vascular de la raíz del ranúnculo (una dicotiledónea).
La endodermis, que está por fuera del periciclo, se considera parte de
la corteza. La endodermis contiene las bandas de Caspary
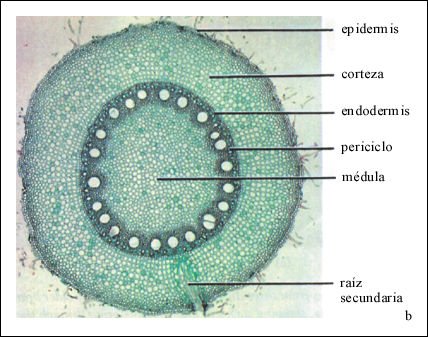
Corte transversal de la raíz de una planta de maíz (una monocotiledónea), que muestra el cilindro vascular que rodea la médula.
Parte de una raíz secundaria que surge del periciclo se puede ver en
la porción inferior de esta microfotografía.
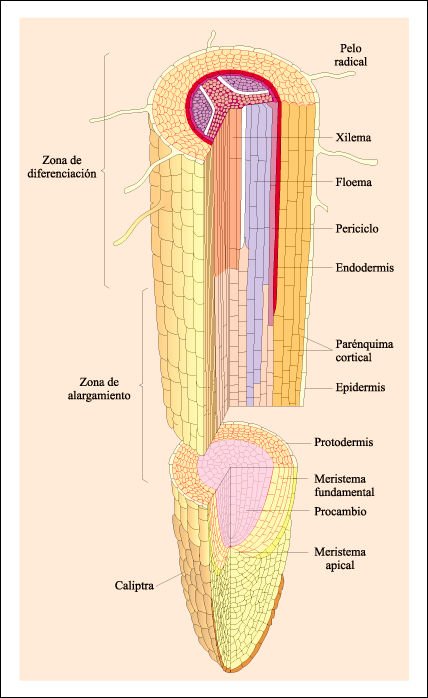
Las regiones de crecimiento de una raíz de dicotiledónea.
Las
células nuevas se producen por división de las células
del meristema apical. Las células que se encuentran por encima del meristema
sufren una serie característica de cambios a medida que aumenta la distancia
entre ellas y el ápice de la raíz. Primero hay una tasa máxima
de división, seguida por alargamiento celular con pocas divisiones posteriores.
Cuando las células se alargan, se diferencian en los tres meristemas
primarios que originan los tres sistemas de tejido de la raíz. La protodermis
se transforma en epidermis, el meristema fundamental, en corteza, y el procambio
se transforma en el xilema primario y el floema primario. Algunas de las células
producidas por el meristema apical se diferencian y forman la caliptra protectora
de la raíz.
Tallos
Los
tallos portan las hojas de la planta y son la vía por la cual las sustancias
se transportan desde las raíces hacia las hojas, y viceversa.
Los tallos verdes, al igual que las hojas, tienen una capa externa de células epidérmicas cubiertas con una cutícula. La masa del tallo joven es tejido fundamental, que puede dividirse en un cilindro externo (la corteza) y un núcleo interno (la médula). El tejido fundamental está compuesto principalmente por células parenquimáticas, pero también puede contener células colenquimáticas y esclerenquimáticas (fibras y esclereidas).
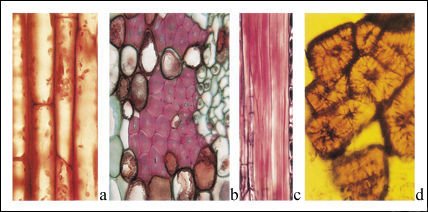
Algunos tipos de células del tejido fundamental de los tallos.
a) Células colenquimáticas vistas en corte longitudinal. b) Corte
transversal y c) corte longitudinal de fibras del floema del tallo de un tilo
(Tilia americana ). Sólo una porción de la longitud de las fibras
puede verse en c). d) Las esclereidas, otro tipo de células esclerenquimáticas,
tienen paredes lignificadas muy gruesas.
Los tejidos vasculares consisten en floema y xilema. En las angiospermas, las células de conducción del floema son los miembros de tubo criboso, células vivas con paredes terminales perforadas, que forman tubos cribosos continuos. Asociada íntimamente con cada miembro de tubo criboso hay una célula acompañante.
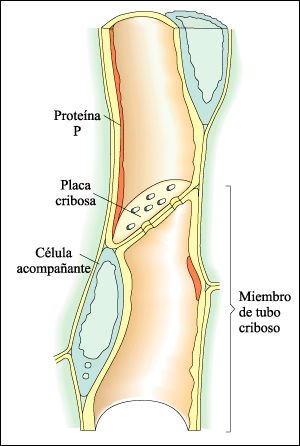
En las angiospermas, los elementos conductores del floema son tubos cribosos, constituidos por células individuales, los miembros de tubo criboso.
Las células de los tubos cribosos, que carecen de núcleo a la
madurez, se encuentran habitualmente en íntima asociación con
las células acompañantes, que tienen núcleo. Los miembros
de tubo cribosos se unen a los otros miembros del tubo por sus extremos mediante
placas cribosas. En la figura se observa una vista longitudinal de tubo criboso
en el tallo del zapallito (Cucurbita maxima). La proteína P tapiza la
superficie interna de las paredes celulares de los miembros de tubo criboso.
El tejido de conducción del xilema está constituido por una serie de traqueidas o vasos. Las traqueidas y los vasos se caracterizan por tener paredes secundarias gruesas y están muertos a la madurez funcional.
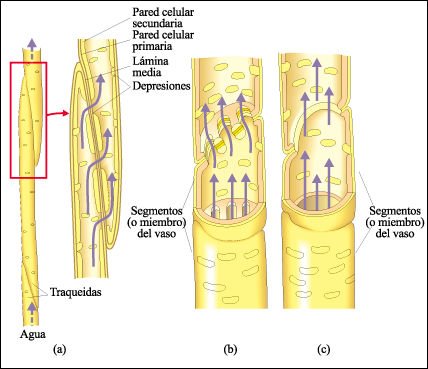
Las traqueidas y los vasos, las células que componen el xilema en las angiospermas.
a) Las traqueidas son un tipo más primitivo y menos eficiente de célula
conductora que predominan en las gimnospermas. El agua que se mueve de una traqueida
a otra pasa a través de depresiones. Estas depresiones no son perforaciones,
sino simplemente áreas en las cuales no hay una pared celular secundaria.
Por lo tanto, el agua que se mueve de una traqueida a otra pasa a través
de dos paredes celulares primarias y de la laminilla media. Los vasos, característicos
de las angiospermas, difieren de las traqueidas en que las paredes primarias
y las laminillas medias de los vasos están perforadas en los extremos,
donde se unen con otros vasos. b) Puede haber numerosas perforaciones en células
contiguas de miembros de vaso, o c) las paredes contiguas pueden disolverse
por completo cuando las células maduran formando una sola abertura. Los
vasos se caracterizan también por ser más cortos y más
anchos que las traqueidas y sus paredes contiguas son menos oblicuas. Los vasos
se conectan con otros vasos y también con otras células por depresiones
de las paredes laterales. d) Vista del interior de un vaso en el xilema de una
raíz de sostén de una planta de maíz
El
floema y el xilema también contienen células parenquimáticas
y fibras.
La altura (o longitud) de las partes aéreas de la planta se incrementa por el crecimiento primario del sistema de vástago. Los primordios foliares surgen del meristema apical del vástago. Cuando los nudos se separan por el alargamiento de los internudos, pequeñas masas de meristema (yemas) se forman en las axilas de las hojas. Estas yemas axilares pueden permanecer latentes o pueden originar ramas o vástagos especializados. Entre los vástagos especializados que pueden formarse están los tubérculos, los tallos rastreros (estolones) y los rizomas, por medio de los cuales muchas especies se reproducen asexualmente, produciendo poblaciones de individuos genéticamente idénticos.
Cortes transversales de dos tallos de dicotiledónea y de un tallo de monoco-tiledónea. a) En la alfalfa, una dicotiledónea, el cilindro vascular está formado por haces vasculares separados.
b) En este tallo joven del tilo, también una dicotiledónea, el tejido vascular forma un cilindro continuo. Este tallo contiene conductos de mucílago, que se tiñen de rojo.

c) En el maíz, una monocotiledónea, numerosos haces vasculares están dispersos en el tejido fundamental.
Crecimiento primario del sistema del vástago
El
sistema del vástago incluye el tallo y todas las estructuras que se desarrollan
a partir de él; típicamente incluye todas las partes aéreas
de la planta.
Primero
ocurre la división celular, luego el alargamiento de las células
y, finalmente, la diferenciación.
Los
primordios foliares surgen del meristema apical del vástago. Cuando los
nudos se separan por el alargamiento de los internudos, pequeñas yemas
se forman en las axilas de las hojas. Estas yemas axilares pueden permanecer
latentes o pueden originar ramas o vástagos especializados. Entre los
vástagos especia-lizados que pueden formarse están los tubérculos,
los estolones y los rizomas, por medio de los cuales muchas especies se reproducen
asexualmente, produciendo poblaciones de individuos genéticamente idénticos.
Crecimiento
secundario
La
mayor parte de las plantas no sólo alcanzan mayor altura con la edad,
sino que también crecen en diámetro. El proceso por el cual las
dicotiledóneas leñosas incrementan el grosor de sus troncos, tallos,
ramas y raíces se conoce como crecimiento secundario. Los así
llamados "tejidos secundarios" no derivan de meristemas apicales sino
que son producidos por meristemas laterales conocidos como cambio vascular y
cambio suberoso.
Este
crecimiento surge primariamente del cambio vascular, una vaina de tejido meristemático
que rodea completamente al xilema y está rodeada completamente por el
floema. Las células de cambio se dividen durante la estación de
crecimiento, añadiendo nuevas células de xilema (xilema secundario)
a sus superficies externas.
A medida que el tronco incrementa su diámetro, la epidermis finalmente se rompe y es reemplazada por corcho.

Etapas de crecimiento de un tallo de dicotiledónea.
a. Tallo de una dicotiledónea antes de que se inicie el crecimiento secundario.
b. Comienzos del crecimiento secundario. El xilema y el floema secundarios son
producidos por el cambio vascular, un tejido meristemático formado en
una etapa tardía del crecimiento pri-mario. A medida que el tronco incrementa
su diámetro, la epidermis se estira y se desgarra. Esto es acompañado
por la formación del cambio suberoso, a partir del cual se forma el cor-cho,
que reemplaza a la epidermis.
c. Corte transversal de un tallo de tres años que muestra los anillos
de crecimiento anuales. Los radios son hileras de células vivas que transportan
nu-trientes y agua lateralmente (a través del tronco). En el perímetro
de la capa más externa de crecimiento del xilema secundario se encuentra
el cambio vascular, rodeado por una banda de floema secundario. El floema primario
y la corteza finalmente se desprenderán. En un tallo más viejo,
el cilindro delgado de floema secundario activo se encuentra en la inmediata
proximidad del cambio vascular. Los tejidos que se encuentran por fuera del
cambio vascular, incluido el floema, constituyen la corteza.
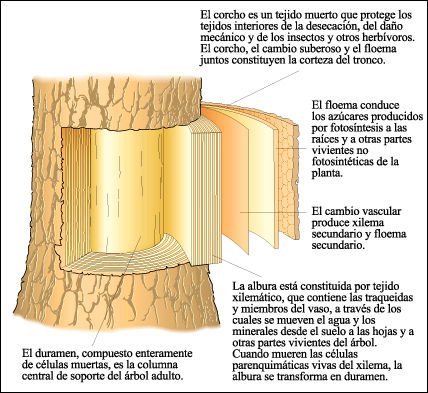
Un tronco de árbol en el que se muestran las relaciones de las capas concéntricas sucesivas.
El duramen está compuesto enteramente de células muertas. La corteza
y la epidermis, que están por fuera del floema en un tallo verde, se
desprenden durante el crecimiento secundario.


